|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
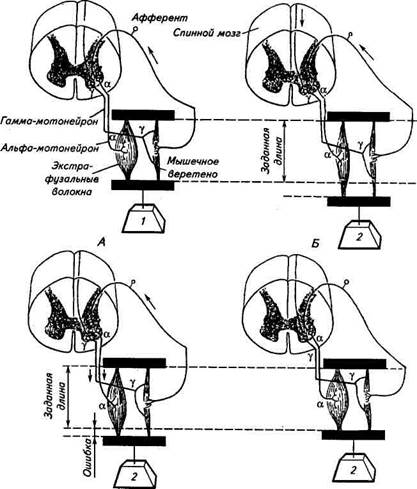
СУХОЖИЛЬНЫЙ РЕЦЕПТОР ГОЛЬДЖИДругие важнейшие афферентные рецепторы двигательного анализатора—сухожильные рецепторы Гольджи (см. рис. 15.2), расположенные главным образом на границе мышечной и сухожильной тканей. Сухожильные рецепторы напоминают по форме мышечные веретена. Функцию вспомогательного аппарата выполняют сухожильные волокна, покрытые соединительнотканной оболочкой — капсулой. Сухожильные волокна внутри капсулы лежат не так плотно, как снаружи, и имеют меньший диаметр. К рецептору подходит одиночное миелинизированное волокно диаметром 8...15мкм, которое внутри капсулы многократно делится, теряя миелиновую оболочку; его тонкие кустиковидные окончания располагаются на сухожильных волокнах. Эфферентных окончаний в отличие от мышечных веретен сухожильные рецепторы Гольджи не имеют. Электрическая реакция рецепторов Гольджи на растяжение следующая: рецепторный потенциал и последовательность потенциалов действия напоминает таковую вторичных афферентных окончаний мышечных веретен (см. рис. 15.3), т. е. сухожильные рецепторы Гольджи являются медленноадаптирующимися рецепторами растяжения — детекторами амплитуды. РЕФЛЕКС НА РАСТЯЖЕНИЕ МЫШЦЫ Рассмотрим некоторые простые примеры функционирования двигательного анализатора с участием мышечных веретен и рецепторов Гольджи. В формировании миотатического рефлекса, или рефлекса на растяжение мышц (рис. 15.5), принимают участие афферентные нейроны, образующие в мышечных веретенах пер-
В г Рис.15.5. Рефлекс на растяжение мышцы: А. В исходном «заданном» состоянии груз небольшой массы (1) удерживается экстра-фузальными волокнами мышцы. В нервных волокнах, образующих афферентные окончания, регистрируются только редкие спонтанные потенциалы действия. Б. При увеличении массы груза (2 > 1) мышца с мышечными веретенами растягивается. В афферентных волокнах повышается частота потенциалов действия, которые поступают через синап-тические контакты на а-мотонейроны (показано стрелкой в направлении от мышечного веретена) и возбуждают их. От а-мотонейронов потенциалы действия направляются к экс-трафузальным мышечным волокнам (стрелки в направлении к мышце) и через синаптичес-кие контакты вызывают сокращение мышцы. В. Сокращение мышцы произошло не до заданной длины. Ликвидация «ошибки» осуществляется с помощью фузимоторных гамма-нейронов, образующих двигательные окончания на интрафузальных мышечных волокнах веретен. Г. Мышца возвращается к заданной длине 41—3389 вичные афферентные окончания, и ос-мотонейроны, которые обеспечивают двигательную иннервацию экстрафузальных мышечных волокон. При растяжении мышцы происходит растяжение и мышечных веретен, что сопровождается повышением частоты потенциалов действия в афферентных волокнах. Поскольку афферентные нейроны синаптически связаны в ЦНС с а-мотонейронами, то в последних также повышается частота потенциалов действия. Распространяясь по эфферентным волокнам, потенциалы действия через синаптические окончания вызывают сокращение — укорочение длины мышцы. Уменьшение растягивающего воздействия на интрафузальные волокна снижает частоту потенциалов действия в афферентных нервных волокнах, и система возвращается в состояние, близкое к исходному. Однако полного восстановления первоначальной длины данная система не обеспечивает. Оставшуюся небольшую разницу между исходной длиной мышцы (до растяжения) и длиной после рефлекторного сокращения (ее называют ошибкой) система не может определить. Для этого потребовалось бы звено обратной связи, т. е. мотонейрон с неограниченно высокой чувствительностью. Способствует возврату мышц к исходной «заданной» длине так называемая фузимоторная система, включающая в себя интрафузальные мышечные волокна и фузимоторные (у) мотонейроны, которые образуют на интрафузальных мышечных волокнах двигательные синапсы. Активация этой системы потенциалами действия из двигательных центров анализатора вызывает сокращение концевых участков веретена и тем самым растяжение центрального несокращаюшегося участка, где находятся афферентные первичные окончания. Это приведет к дополнительному увеличению частоты потенциалов действия в афферентном нейроне, которое будет восприниматься а-мотонейро-ном с последующей посылкой к синаптическим окончаниям экстрафузальных волокон эфферентных потенциалов действия. В результате этого в мышце произойдет дополнительное сокращение и исходная длина будет достигнута. Из вышеизложенного становится ясно, что миотатический рефлекс служит для поддержания постоянной длины мышцы при изменениях воздействующей на нее нагрузки. Этот механизм у животных, как, по-видимому, и у человека, осуществляется без контроля сознания и играет определяющую роль в поддержании позы. Ответственные за положение тела в пространстве мышцы-разгибатели должны иметь определенную заданную длину и в противовес силы тяжести удерживать конечности животного в выпрямленном состоянии. Сухожильные рецепторы Гольджи соединены с мышечными волокнами последовательно (а не параллельно, как мышечные веретена), поэтому они должны реагировать на изменение напряжения мышцы, а не длины. Было обнаружено, что через тормозные интернейроны афферентная импульсация i рсигишрмп I <>iii.*i.*m оказывает влияние на а-мотонейроны, снижая vi>">'< m, us,п> ши ности. Это, например, может проявляться в понижпни... ими потенциалов действия, посылаемых к синапсам >kci|),h|>v ини.мы* мышечных волокон, что предотвращает чрезмерное пшфнжгнж мышцы. Предполагается также, что сигнализация сухожильными рецепторами о мышечном напряжении а-мотонейронам cnoioit ствует исправлению неточностей в регуляции длины мышц мин татическими рефлексами. Дата добавления: 2015-12-15 | Просмотры: 1002 | Нарушение авторских прав |