|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
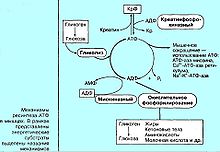
Анаэробное и аэробное энергообразование в тканях человекаНекоторые ткани животных и человека отличаются повышенной устойчивостью к гипоксии (особенно мышечная ткань). В обычных условиях синтез АТФ идет аэробным путем, а при напряженной мышечной деятельности, когда доставка кислорода к мышцам затруднена, в состоянии гипоксии, а также при воспалительных реакциях в тканях доминируют анаэробные механизмы регенерации АТФ. В скелетных мышцах выявлены 3 вида анаэробных и только один аэробный путь регенерации АТФ.
3 вида анаэробного пути синтеза АТФ К анаэробным относятся: Креатинфосфатазный (фосфогеный или алактатный) механизм — перефосфорилирование между креатинфосфатом и АДФ Миокиназный — синтез (иначе ресинтез) АТФ при реакции трансфосфорилирования 2 молекул АДФ(аденилатциклаза) Гликолитический — анаэробное расщепление глюкозы крови или запаса гликогена, заканчивающийся образованием молочной кислоты (иначе именуется «лактатным»). Необходимо отметить, что прямым следствием гликолиза является критическое снижение рН тканей — ацидоз. Это ведет к снижению эффективного транспорта кислорода гемоглобином, и формирует положительную обратная связь. Каждый механизм имеет свое время удержания максимальной мощности и оптимум энергообеспечения тканей. Наибольшая мощность и наименьшее время удержания: креатинфосфаткиназный механизм (3600 (Дж·кг)/мин, при времени 6—12 сек) лактатный (2510 (Дж·кг)/мин, при времени 30—60 сек) аэробный (600 (Дж·кг)/мин, при времени около 600 секунд). [править] Примечания ↑ Газогенерирующие контейнерные системы GasPak: Инструкция МК. — OOO "МК, официальный дистрибьютер Becton Dickinson International", 2010. — С. 7. ↑ 1 2 3 К.Д.Пяткин Микробиология с вирусологией и иммунологией. — М:"Медицина", 1971. — С. 56. ↑ Л.Б.Борисов Медицинская микробиология, вирусология и иммунология. — МИА, 2005. — С. 154-156. — ISBN 5-89481-278-X ↑ Д.Г.Кнорре Биологическая химия:Учеб. для хим., биол. и мед.спец.вузов. — 3. — М.:Высшая школа, 2000. — С. 134. — ISBN 5-06-003720-7 ↑ D.A.Eschenbach, P.R.Davick, B.L.Williams Prevalence of hydrogen peroxide-producing Lactobacillus species in normal women and women with bacterial vaginosis.. — J Clin Microbiol. 1989 February; 27(2): 251–256.. ↑ М.В.Гусев,Л.А.Минеева Микробиология. — М:МГУ, 1992. — С. 56. ↑ А.А. Воробьев Атлас по медицинской микробиологии, вирусологии и иммунологии. — МИА, 2003. — С. 44. — ISBN 5-89481-136-8 ↑ Л.Б.Борисов Руководство к лабораторным занятиям по медицинской микробиологии, вирусологии и иммунологии. — Медицина, 1992. — С. 31-44. — ISBN 5-2225-00897-6 ↑ J.H.Brewer, D.L.Allgeier Disposable hydrogen generator.. — Science 147:1033-1034.. — 1966. ↑ J.H.Brewer, D.L.Allgeier Safe self-contained carbon dioxide-hydrogen anaerobic system.. — Appl. Microbiol.16:848-850.. — 1966. ↑ G.F.Smirnova Metabolism peculiarities of bacteria restoring chlorates and perchlorates.. — Microbiol Z. 2010 Jul-Aug;72(4):22-8.. ↑ Филиппович Ю.Б., Коничев А.С., Севастьянова Г.А. Биохимические основы жизнедеятельности организма человека. — Владос, 2005. — С. 302. — ISBN 5-691-00505-7 [править] См. также Аэробы [править] Ссылки Анаэробии (анаэробы) // Энциклопедический словарь Брокгауза и Ефрона: В 86 томах (82 т. и 4 доп.). — СПб.: 1890—1907. Анаэробы // Энциклопедический словарь Брокгауза и Ефрона: В 86 томах (82 т. и 4 доп.). — СПб.: 1890—1907.
Питание бактерий. Питательные вещества – источники углерода и азота. Классификация бактерий по типам питания Аутотрофы и хемоорганотрофы Катаболи́зм (от греч. καταβολή, «основание, основа») — процесс метаболического распада, разложения на более простые вещества или окисления какого-либо вещества, обычно протекающий с высвобождением энергии в виде тепла и в виде АТФ. Катаболические реакции лежат в основе диссимиляции: утраты сложными веществами своей специфичности для данного организма в результате распада до более простых. Анаболи́зм (от греч. ἀναβολή, «подъём») — совокупность химических процессов, составляющих одну из сторон обмена веществ в организме, направленных на образование составных частей клеток и тканей. Анаболизм взаимосвязан с противоположным процессом — катаболизмом, так как продукты распада различных соединений могут вновь использоваться при анаболизме, образуя в иных сочетаниях новые вещества. Процессы анаболизма, происходящие в зелёных растениях с поглощением энергии солнечных лучей (см. Фотосинтез), имеют планетарное значение, играя решающую роль в синтезе органических веществ из неорганических. Анаболизм (пластический обмен, ассимиляция) — одна из сторон обмена веществ. Включает процессы
По типу питания все организмы делятся на автотрофов и гетеротрофов. Автотрофы, что в переводе с греческого означает «самопитающиеся», могут строить все соединения своих клеток из углекислоты и других неорганических веществ. Источником энергии для них служит либо свет (фотоавтотрофы), либо они ее получают при окислении минеральных соединений (хемоавтотрофы). Таким образом, ни для конструктивных, ни для энергетических процессов органические субстраты автотрофам не требуются.
Гетеротрофы также могут усваивать углекислоту. Однако им необходимы органические соединения как основные источники углерода, а в большинстве случаев и субстраты для получения энергии (хемоорганогетеротрофы). Такой тип питания реализуется у человека, животных и многих микроорганизмов. Лишь для некоторых бактерий, нуждающихся в готовых органических соединениях, источником энергии является свет (фотогетеротрофы).
Среди автотрофов наиболее широко распространены организмы, использующие лучистую энергию. Они представлены высшими растениями, водорослями и рядом бактерий, способных к фотосинтезу.
Хемоавтотрофы обнаружены только среди бактерий, т. е. только среди прокариотных организмов, причем количество их сравнительно невелико. Однако по своим физиолого-биохимическим свойствам, геохимической деятельности и значению для некоторых областей народного хозяйства эти микроорганизмы весьма интересны.
Существование хемоавтотрофов было открыто С. Н. Виноградским. Началом послужили его работы (1885—1889) по изучению нитчатых микроорганизмов, называемых серобактериями (Beggiatoa) и железобактериями (Leptothrix ochracea). В результате проведенных наблюдений Виноградский пришел к выводу, что жизнедеятельность указанных форм связана с окислением соответственно сероводорода и серы до серной кислоты или закисного железа в окисное и оба процесса имеют энергетическое значение.
Свою гипотезу Виноградский блестяще доказал, выделив (1890—1892) чистые культуры нитрифицирующих бактерий (Nitrosomonas и Nitrobacter), которые росли на минеральных средах, окисляя аммонийный азот или нитриты и фиксируя при этом углекислоту.
В настоящее время хемоавтотрофов подразделяют на следующие группы, получившие свои названия соответственно природе окисляемых субстратов:
1. Нитрифицирующие бактерии.
2. Водородные бактерии.
3. Серобактерии и тионовые бактерии.
4. Железобактерии.
Кроме того, к хемоавтотрофам, видимо, принадлежит недавно обнаруженный Н. Н. Ляликовой-Медведевой микроорганизм Stibiobacter, окисляющий окислы трехвалентной сурьмы (Sb2О3) до пятивалентной (Sb205).
Таким образом, выявлены хемоавтотрофы, способные получать энергию в результате окисления минеральных соединений пяти элементов: Н, N, S, Fe и Sb.
Факторы роста и их источники. Источники минеральных элементов. Способы и механизмы переноса питательных веществ через мембрану. Энергетические потребности бактерий. Пути получения энергии у аутотрофов (фотосинтез, хемосинтез). Источники и пути получения энергии у хемоорганотрофов. Аэробный и анаэробный типы биологического окисления у бактерий. Аэробные, анаэробные, факультативно анаэробные и микроаэрофильные бактерии. Способы создания анаэробных условий. Задачи, этапы, преимущества и недостатки бактериологического (культурального) метода исследования. Рост и размножение микроорганизмов. Способы размножения. Бинарное (простое) деление, механизм. Размножение бактериальных популяций. Принципы и методы культивирования бактерий. Питательные потребности микробов. Питательные среды для культивирования бактерий. Требования к питательным средам. Классификация питательных сред. Условия и техника культивирования бактерий. Техника посева на питательные среды. Закономерности и характер роста бактерий на плотных и жидких питательных средах. Способы выделения чистых культур аэробных и анаэробных бактерий. Свойства, используемые для идентификации выделенных культур.
Дата добавления: 2015-12-15 | Просмотры: 1512 | Нарушение авторских прав |
 ПЕРЕЧЕНЬ КОНТРОЛЬНЫХ ВОПРОСОВ
ПЕРЕЧЕНЬ КОНТРОЛЬНЫХ ВОПРОСОВ