|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Немембранные структуры клеткиРибосома (рис.15) представляет собой элементарную клеточную машину синтеза любых белков клетки. У всех растительных (да и животных) организмов они идентичны. Их размеры варьируют от 17 до 23 нм в диаметре, вследствие чего они не видны в световой микроскоп. В состав эукариотической рибосомы входят четыре молекулы РНК, различающиеся по длине: от 120 до 4700 нуклеотидов, которые связаны с белком. Число рибосом в клетке, как правило, чрезвычайно велико (до 107 шт.). Рибосомы могут образовывать комплексы, называемые полирибосомами или полисомами. Клетки, активно синтезирующие белки, имеют, как правило, много полисом.
Рисунок 15. Внешний вид рибосомы, вид сверху и сбоку. Задание 9. Микрофотография гранулярного эндоплазматического ретикулума
Рассмотреть микрофотографии гранулярного эндоплазматического ретикулума (рис. 165, с. 281, Ченцов 2004) и зарисовать. Обозначить на рисунке эндоплазматический ретикулум, рибосомы.
Задание 9. Микрофотографии молодой растительной клетки
Рассмотреть микрофотографии (рис. 1-2, с.14, Атлас по биологии клетки,) и зарисовать. Обозначить на рисунке органеллы клетки и рибосомы. РАБОТА № 5.
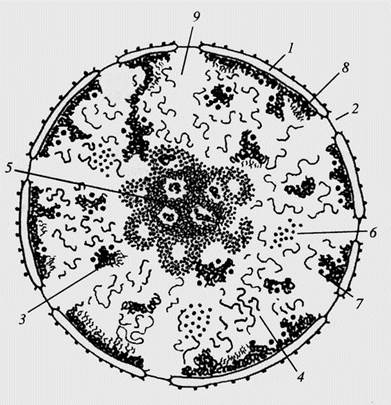
Ядро - это органоид, в котором сосредоточена основная масса генетического материала клетки. Важнейшим внутриядерным процессом является воспроизводство самого генетического материала, необходимое для последующего деления клетки или для интенсификации ее синтетической активности. Реализация генетической информации заключается в целой цепи синтетических процессов и имеет своей конечной целью синтез определенных белков, обусловливающих жизнедеятельность клетки. Начальные этапы реализации генетической информации происходят именно в ядре, важнейшая функция которого состоит в производстве всех РНК, обеспечивающих синтез белка. В настоящее время в ядре различают следующие компоненты: ядерная оболочка, хроматин (хромосомный материал), ядрышко и ядерный сок (кариоплазма) (рис. 16).
Рисунок 16. Схема строения клеточного ядра. 1 – ядерная оболочка (две мембраны и перинуклеарное пространство); 2 – ядерная пора; 3 – конденсированный хроматин; 4 – диффузный хроматин; 5 – ядрышко (гранулярный и фибриллярный компонент, в светлых зонах находится рДНК); 6 – интерхроматиновые гранулы; 7 – перихроматиновые гранулы; 8 – перихроматиновые фибриллы; 9 – кариоплазма.
Эти компоненты отличаются друг от друга по своему биохимическому составу и ультраструктуре, так как несут различные функциональные нагрузки. Необходимо еще раз подчеркнуть, что, несмотря на эти различия, работа всех компонентов ядра направлена на выполнение генетической функции хромосомного материала — репликацию ДНК и синтез РНК. Ядро выполняет две важнейшие функции: контроль жизнедеятельности клеток и хранение и передача генетической информации. Ядро содержит нуклеиновые кислоты - ДНК и РНК, белки, небольшие молекулы и ионы. Почти у всех растительных клеток оно имеет сферическую форму и состоит из нуклеоплазмы, хроматина и ядерной оболочки. Хроматином называют комплексы ДНК с белками. У растений ядро клетки включает также небольшое сферическое тельце, получившее название ядрышка, которое представляет собой область внутри ядра, где локализованы гены, ответственные за синтез рибосомной РНК. У некоторых видов растений в ядрах клеток имеется несколько ядрышек. Участки хромосом, в которых происходит формирование ядрышка, называются ядрышковыми организаторами или ядрышко организующими районами (ЯОР). Хроматин состоит из комплекса, образуемого молекулами ДНК и белками основной природы - гистонами. Кроме них в состав хромосом могут входить высокомолекулярные белки и РНК. Диаметр ядра у высших организмов обычно колеблется от 10до 30 мкм. Ядерная оболочка является дифференцированной частью эндоплазматической сети. Оболочка состоит из двух мембран, разделенных перинуклеарным пространством. Ядерная оболочка пронизана порами, которые контролируют обмен между ядром и цитоплазмой. Через поры проходят мелкие молекулы, ионы, а также макромолекулы, в частности такие, как рибонуклеопротеиды. В ядрышках протекает синтез рибосомных РНК. Ядрышки часто прилегают к той части хроматина, где они образуются. Их размеры и в некоторых случаях количество характеризуют повышенную степень активности клетки (синтез белков).
Задание 1. Ультраструктура интерфазных ядер
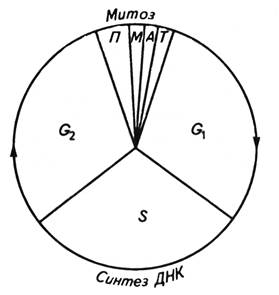
Рассмотреть микрофотографии интерфазных ядер (рис. 29, 30, с. 70-71, Ченцов 2004,) и зарисовать. Обозначить на рисунке ядерную оболочку, ядрышко, нуклеоплазму, диффузный хроматин. Задание 2. Животная клетка в период дифференциации Рассмотреть микрофотографию животной клетки в период дифференциации (рис. 1-3, с. 15, Атлас по биологии клетки, 1978) и зарисовать. Обозначить на рисунке клеточную мембрану, ядро, ядерную оболочку, ядрышко, нуклеоплазму, хроматин, ядерные поры. Задание 3. Ультраструктура ядерной мембраны Рассмотреть микрофотографии ядерной мембраны (рис. 8-3, 8-4, 8-6, с. 75, Атлас по биологии клетки, 1978) и зарисовать. Обозначить на рисунке ядро, цитоплазму, внутреннюю ядерную мембрану, наружную ядерную мембрану, перинуклеарное пространство, ядерные поры, хроматин, рибосомы. Задание 4. Ядра растительных клеток. Корешок лука Ядра растительных клеток по своей структуре отличаются от ядер клеток животных несколько иной упаковкой в них хроматина. Именно на это должны обратить внимание студенты, рассматривая светомикроскопический препарат и электронограмму растительной клетки. В качестве объекта для наблюдения можно использовать корешки лука или корешки проростков его семян. Под малым увеличением микроскопа в кончике корня можно видеть чехлик, состоящий из многоугольных уплотненных клеток, и меристему, представленную столбцами клеток в основном прямоугольных или близких к прямоугольным очертаний. Интересно, что клетки одного ряда представляют потомство одной клетки. Форма и структура ядер в меристематических клетках и клетках чехлика различны. Под иммерсионным объективом в центре меристематической клетки видно крупное шаровидное или слегка вытянутое ядро с одним или двумя большими сферическими ядрышками. В отличие от ядер многих клеток животных организмов ядра клеток растений часто имеют в своём составе так называемую хроматиновую сеть. Она представляет собой скопление тонких нитей, очень плотных и интенсивно красящихся.. Видны глыбки и зерна хроматина разных размеров. Крупные глыбки называют хромоцентрами. Некоторые хромоцентры располагаются вблизи ядрышка. Деление растительной клетки начинается с деления ядра. В соматической клетке оно носит название митоза- непрямого или кариокинетического деления, который протекает в меристематических тканях. В результате этого деления из одной клетки образуются две дочерние, имеющие такое же число хромосом, какое было у родительской клетки. Период от окончания одного митоза до окончания следующего получил название клеточного цикла (рис.17). Между двумя клеточными делениями проходит период, во время которого внешне клетка находится в стоянии покоя - стадия интерфазы. Однако этот покой только видимый, а в клетке интенсивно протекают процессы на молекулярном уровне, подготавливающие вновь образовавшиеся клетки к новому делению. Интерфаза включает пресинтетический период (G1), синтетический период (S), постсинтетический период (G2). В период G1 продолжается рост клеток, синтезируются специфические белки и нуклеотиды, необходимые для синтеза ДНК. Период S характеризуется синтезом ДНК (ее количество в клетке удваивается) и гистонов. Удвоение содержания ДНК связано с репликацией хромосом. В конце этого периода каждая из хромосом состоит из двух хроматид.
Рис.17. Схема периодов клеточного цикла. G1 – пресинтетический период, S – синтетический период, G2 – постсинтетический период.
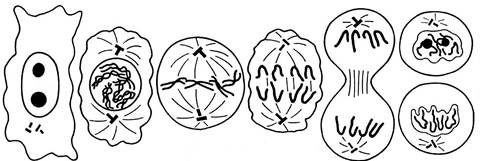
Период G2характеризуется накоплением веществ и энергии, необходимых для протекания митоза. В этот период начинаются процессы конденсации хромосом. Перед расхождением в дочерние клетки хромосомы постепенно переходят в метаболически неактивное состояние. Продолжительность клеточного цикла очень сильно варьирует в зависимости от ткани, вида растения и внешних условий. Например, по данным К. Свенсона и П. Уэбстера, в кончиках боковых корешков конских бобов (Vicia faba) средняя продолжительность клеточного цикла в меристематических клетках при 22 оС составила 14 ч: период G1 занимал 2,5 ч, период S—6 ч, период G2 — 3,5 ч и собственно митоз продолжался 2 ч. Внутри клеточного цикла наиболее сильно варьирует продолжительность периода G1, который может практически отсутствовать в активно делящихся клетках или казаться практически постоянным в случае специализированных тканей. В случае последних часто используют характеристику Go, т. е. неактивной фазы G1. В митозе выделяют два взаимосвязанных процесса - кариокинез (деление ядра) и цитокинез (деление цитоплазмы). Митоз состоит из следующих стадий: профазы, метафазы, анафазы и телофазы (рис. 18). Профаза характеризуется продолжением процесса конденсации хроматина, в результате чего хромосомы становятся видимыми в световой микроскоп. На этой стадии исчезает ядрышко (ядрышки). В конце профазы появляются микротрубочки и начинает формироваться веретено деления. Затем клетка плавно переходит в метафазу. К началу метафазы ядерная оболочка разрушается, а хромосомы достигают максимального уровня конденсации.
1 2 3 4 5 6 Рис. 18. Схема митотического деления клетки. 1 – интерфаза; 2 – профаза; 3 – метафаза; 4 – анафаза; 5 - ранняя телофаза; 6 - поздняя телофаза. В то же время окончательно формируется веретено деления, состоящее из пучков микротрубочек: опорных - идущих от полюса к полюсу, и тянущих - от полюсов к центромерам дихроматидных хромосом. Хромосомы выстраиваются перпендикулярно к нитям веретена на равном удалении от полюсов, образуя метафазную пластинку. В анафазе центромеры делятся в продольном направлении и хроматиды (теперь это самостоятельные хромосомы) под действием тянущих нитей веретена начинают движение к полюсам. Деление центромер происходит синхронно за счет разделения белков когезинов. К концу анафазы в экваториальной плоскости клетки на опорных нитях веретена образуются небольшие узелки, которые в дальнейшем (по завершению телофазы) сливаются и дают начало первичной клеточной перегородке. Заключительная фаза митоза - телофаза. Во время этой фазы начинается деконденсация хромосом, восстанавливаются ядрышки и формируется ядерная оболочка, начинает закладываться клеточная перегородка. Митоз завершается формированием клеточной стенки, а два дочерних ядра в двух новых клетках вступают в интерфазу. Митоз свойствен всем эукариотам. Его биологическое значение заключается в том, что в результате обе дочерние клетки имеют одинаковое с родительской число хромосом. Помимо митоза имеют место еще три типа деления ядра соматических клеток: амитоз, эндомитоз и политения. Амитоз - это прямое деление ядра, при котором оно делится перетяжкой на две части. Затем происходит разделение цитоплазмы клетки и возникает клеточная перегородка. Амитотическое деление приводит к неравномерному распределению ДНК в дочерних клетках. Амитоз, как правило, свойствен клеткам высокополиплоидных дифференцированных тканей, таких, как клетки стенок завязи, крахмалообразующие клетки клубней картофеля, клетки перисперма и др. Эндомитоз. При этом типе деления (рис. 19) ядерная оболочка не распадается. Удвоение хромосом, как и при митозе, происходит во время предшествующей интерфазы. Процесс удвоения проходит неоднократно, поэтому число хромосом в ядре и размеры самого ядра увеличиваются. Эндомитоз впервые был обнаружен в клетках тапетума шпината (Spinacia sativa), а затем в антиподах семейств астровых (Asteraceae) и лютиковых (Ranunculaсеае). При эндомитозе хромосомы проходят те же стадии, что и при нормальном митозе. Встречаются два типа этого деления, отличающиеся тем, что в одном случае хроматиды в эндоанафазе расходятся, а в другом - нет. Последний приводит к политении.
Рис.19. Схема эндомитоза в клетках тапетума Spinacia oleracea: А — эндопрофаза; Б — эвдометафаза; В— эндоанафаза; Г— эндотелофаза.
Политения. Ее можно рассматривать как частный случай эндомитоза. При политении образуются гигантские хромосомы за счет многократной редупликации хроматид без разделения центромеры. При этом степень конденсации хроматид меньше, чем у митотических хромосом. Хроматиды плотно прилегают друг к другу, и хромомеры многочисленных хроматид образуют поперечные диски и пуффы. Впервые политенные хромосомы были обнаружены в слюнных железах личинки комара, а затем и в ядрах эндосперма, синергид и антипод представителей различных семейств растений. Задание 5. Митоз в клетках корешка лука
Деления клеток происходят только в зоне деления. При большом увеличении микроскопа в ней можно найти как неделящиеся клетки (стадия интерфазы), так и клетки во всех стадиях митоза. В интерфазе клетки имеют прямоугольные очертания, окружены хорошо заметной оболочкой. Ядра округлые и овальные, в них видны 1 - 2 окрашенных в черный цвет округлых, крупных ядрышка и мелкие глыбки хроматина. В профазе в ядре заметны более крупные глыбки хроматина. Затем, в результате дальнейшей конденсации хромосом, в ядре появляется клубок вначале тонких, а позднее более толстых и коротких нитей. В ранней профазе еще хорошо заметны ядрышки и ядерная оболочка, а к концу профазы ядрышко и ядерная оболочка исчезают. Хромосомы в виде коротких нитей оказываются лежащими в центральной части цитоплазмы. Для следующей стадии деления - метафазы характерны два процесса: 1) завершение образования (начавшегося еще в профазе) митотического аппарата деления, состоящего из тонких нитей, тянущихся от одного полюса клетки к другому. Следует обратить внимание на то, что в клетках высших растений митотический аппарат деления формируется без участия клеточного центра; 2) перемещение хромосом к центру клетки (метакинез) и расположение их в виде экваториальной пластинки. При этом часть нитей веретена (так называемые прикрепленные нити) оканчиваются на лежащих в центральной области хромосом - кинетохорах. Остальные нити веретена деления идут через всю клетку от одного ее полюса к другому. Хромосомы в экваториальной пластинке располагаются центромерными участками друг к другу, а концы их обращены наружу. Поэтому при рассмотрении их сверху они образуют фигуру, напоминающую звезду (стадия материнской звезды). В метафазе каждая хромосома состоит из двух хроматид (сестринских хромосом). Следует помнить, что удвоение хромосом происходит в интерфазе в синтетическом S-периоде. Условно концом метафазы можно считать тот момент, когда хроматиды начинают отходить друг от друга и соединены лишь в области центромеров. В анафазе митоза обеспечивается равномерное распределение генетического материала по дочерним клеткам. В ранней анафазе хромосомы повернуты центромерами к полюсам клетки, а концы их обращены к центру клетки. В поздней анафазе хромосомы уже собираются на полюсах клетки. Расхождение хромосом происходит очень быстро. В телофазе хромосомы начинают деконденсироваться, становится заметным матрикс хромосом. Появляется ядерная оболочка и восстанавливаются ядрышки. Одновременно в поздней анафазе и ранней телофазе в центре клетки в области веретена начинает образовываться перегородка, которая растет от центра клетки к периферии и делит клетку на две дочерние клетки. Рассмотреть препарат, найти и зарисовать клетки корешка лука со стадиями митоза соответственно схеме митоза (рис. 18). Обозначить на рисунке стадии митоза, ядро, хроматин, хромосомы, веретено деления, цитокинез. Задание 6. Митоз в животной клетке
Рассмотреть при иммерсионном увеличении микроскопа препаратспермогония речного рака. Найти и зарисовать стадии митоза животной клетки. Отметить отличия цитокинеза раститальной и животной клетки. В интерфазе в сперматогониях видны округлые ядра с 1-2 ядрышками, глыбками и нитями хроматина (рис. 39, с. 85, Ченцов Ю.С., 1985). В профазе хромосомы начинают конденсироваться и становятся видны в ядре в виде крупных, темноокрашенных глыбок, заполняющих все ядро. В ранней профазе имеются ядрышко и ядерная оболочка. В поздней профазе ядрышко растворяется, а затем разрушается ядерная оболочка. В метафазе заканчивается формирование митотического аппарата деления, образование которого начинается еще в профазе. На препарате на стадии метафазы ахроматиновое веретено деления имеет вид бочонка, состоящего из тонких, темноокрашенных нитей (микротрубочек). Концы нитей собираются на полюсах клетки. В этих местах лежат круглые, окрашенные в черный цвет гранулы - центриоли. В клетках животных в отличие от клеток высших растений клеточный центр принимает участие и в образовании митотического аппарата деления (ахроматинового веретена). Хромосомы, имеющие форму коротких палочек, многочисленные, в начале метафазы перемещаются к центру клетки и здесь хромосомы располагаются по экватору веретена и образуют широкую экваториальную пластинку, в которой трудно различить отдельные хромосомы. Часть ахроматиновых нитей веретена прикрепляется к кинетохорам хромосом в центромерном участке. На следующей стадии - анафазе происходит расхождение хроматид (сестринских хромосом), образующих метафазную хромосому. Следует помнить, что удвоение ДНК и соответственно хромосом происходит в синтетическом (S) периоде интерфазы, а в анафазе осуществляется расхождение хромосом к полюсам клетки. В этой фазе обеспечивается равномерное распределение генетического материала по дочерним клеткам. На препарате можно найти как раннюю, так и позднюю анафазу. Они отличаются лишь расстоянием хромосом от экватора клетки. В поздней анафазе начинает делиться тело клетки. По периферии клетки образуется перетяжка, борозда, которая постепенно углубляется и в дальнейшем делит клетку на две части. В последней стадии деления - телофазе идет реконструкция ядер дочерних клеток. Хромосомы деспирализуются (деконденсируются), нити его слипаются, бледнеют и веретено постепенно исчезает. Следует обратить внимание на то, что деление тела животной клетки идет с периферии, в то время как в клетках высших растений деление тела клетки идет от центра, где образуется пластинка, растущая от центра к периферии. Задание 7. Амитоз эпителиальных клеток
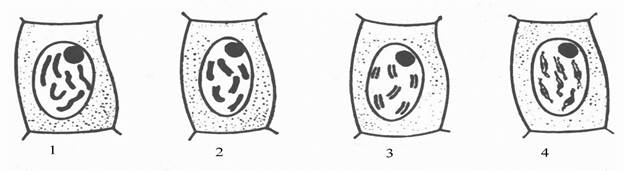
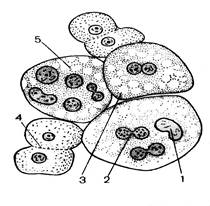
При малом увеличении видны различной величины и формы ядра эпителиальных клеток слизистой оболочки мочевого пузыря. Слабо оксифильная цитоплазма этих клеток почти неразличима, однако с опущенным конденсором и прикрытой диафрагмой можно видеть клетки различной величины. В крупных, неправильной формы округлых клетках содержится одно, два, три и более ядер (рис. 20). Среди неделящихся ядер можно видеть также ядра, находящиеся на разных стадиях амитотического деления. Эти ядра надо изучить при большом увеличении. Во время прямого деления ядро остается в интерфазном состоянии и клетка не перестает функционировать. О начале амитоза свидетельствуют изменения формы ядра. Оно вытягивается в длину, в средней части образуется перетяжка, которая быстро истончается и разрывается. Клетка становиться двуядерной. В дальнейшем может произойти цитотомия. Нередко цитотомия задерживается или вообще не наступает, в результате чего возникают многоядерные клетки. Иногда видно деления ядрышка, которое удлиняется и перешнуровывается.
Рис. 20. Амитоз в клетках эпителия. 1 – ядро; 2- перетяжка; 3 - двуядерная клетка; 4 – цитотомия; 5 – многоядерные клетки.
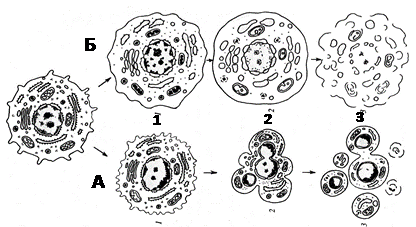
Прямое деление клеток эпителия мочевого пузыря является дегенеративной формой, так как предшествует отмиранию этих клеток. Гибель отдельных клеток или целых групп постоянно встречается у многоклеточных организмов. Причины гибели, процессы морфологического и биохимического характера развития клеточной смерти могут быть различны. Но они четко делятся на две категории: некроз и апоптоз (рис. 21). Некроз связан с нарушением внутреннего гомеостаза в результате нарушения проницаемости клеточных мембран, с необратимыми изменениями митохондрий. Апоптоз – гибель клеток без их химического и физического повреждения на всех стадиях онтогенеза. Это процесс – приводящий не к лизису, не к растворению клеток, а к их фрагментации, распаду. Биологическая роль – удаление отработавших или ненужных на данном этапе клеток, а так же удаление измененных или патологических клеток, особенно мутантных или зараженных вирусами.
Рисунок 21. Два пути клеточной гибели. А – апоптоз: 1 – специфическое сжатие клетки и конденсация хроматина; 2 – фрагментация ядра; 3 – фрагментация тела клетки на ряд апоптических телец. Б – некроз: 1 – набухание клетки, вакуолярных компонентов, конденсация хроматина (кариорексис); 2 – набухание мембранных органоидов, лизис хроматина ядра (кариолизис); 3 – разрыв мембранных компонентов клетки – лизис.
Дата добавления: 2015-12-16 | Просмотры: 1621 | Нарушение авторских прав |