|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Газообмен и транспорт О2
Транспорт О2 осуществляется в физически растворенном и химически связанном виде. Физические процессы, т. е. растворение газа, не могут обеспечить запросы организма в О2. Подсчитано, что физически растворенный О2 может поддерживать нормальное потребление О2 в организме (250 мл*мин-1), если минутный объем кровообращения составит примерно 83 л*мин-1 в покое. Наиболее оптимальным является механизм транспорта О2 в химически связанном виде.
Согласно закону Фика, газообмен О2 между альвеолярным воздухом и кровью происходит благодаря наличию концентрационного градиента О2 между этими средами. В альвеолах легких парциальное давление О2 составляет 13,3 кПа, или 100 мм рт.ст., а в притекающей к легким венозной крови парциальное напряжение О2 составляет примерно 5,3 кПа, или 40 мм рт.ст. Давление газов в воде или в тканях организма обозначают термином «напряжение газов» и обозначают символами Ро2, Рсo2. Градиент О2 на альвеолярно-капиллярной мембране, равный в среднем 60 мм рт.ст., является одним из важнейших, но не единственным, согласно закону Фика, факторов начальной стадии диффузии этого газа из альвеол в кровь.
Транспорт О2 начинается в капиллярах легких после его химического связывания с гемоглобином.
Гемоглобин (Нb) способен избирательно связывать О2 и образовывать оксигемоглобин (НbО2) в зоне высокой концентрации О2 в легких и освобождать молекулярный О2 в области пониженного содержания О2 в тканях. При этом свойства гемоглобина не изменяются и он может выполнять свою функцию на протяжении длительного времени.
Гемоглобин переносит О2 от легких к тканям. Эта функция зависит от двух свойств гемоглобина: 1) способности изменяться от восстановленной формы, которая называется дезоксигемоглобином, до окисленной (Нb + О2 à НbО2) с высокой скоростью (полупериод 0,01 с и менее) при нормальном Рог в альвеолярном воздухе; 2) способности отдавать О2 в тканях (НbО2 à Нb + О2) в зависимости от метаболических потребностей клеток организма.
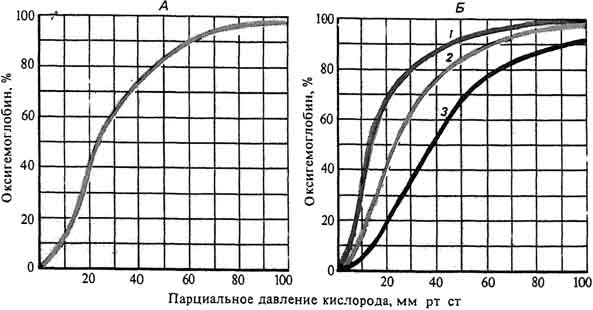
Зависимость степени оксигенации гемоглобина от парциального давления Ог в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина, или сатурационной кривой (рис. 8.7). Плато кривой диссоциации характерно для насыщенной О2 (сатурированной) артериальной крови, а крутая нисходящая часть кривой — венозной, или десатурированной, крови в тканях.
На сродство кислорода к гемоглобину влияют различные метаболические факторы, что выражается в виде смещения кривой диссоциации влево или вправо. Сродство гемоглобина к кислороду регулируется важнейшими факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата. Величина рН и содержание СО2 в любой части организма закономерно изменяют сродство гемоглобина к О2: уменьшение рН крови вызывает сдвиг кривой диссоциации соответственно вправо (уменьшается сродство гемоглобина к О2), а увеличение рН крови — сдвиг кривой диссоциации влево (повышается сродство гемоглобина к О2) (см. рис. 8.7, А). Например, рН в эритроцитах на 0,2 единицы ниже, чем в плазме крови. В тканях вследствие повышенного содержания СО2 рН также меньше, чем в плазме крови. Влияние рН на кривую диссоциации оксигемоглобина называется «эффектом Бора».
Рост температуры уменьшает сродство гемоглобина к О2. В работающих мышцах увеличение температуры способствует освобождению О2. Уменьшение температуры тканей или содержания 2,3-дифосфоглицерата вызывает сдвиг влево кривой диссоциации оксигемоглобина (см. рис. 8.7, Б).
Метаболические факторы являются основными регуляторами связывания О2 с гемоглобином в капиллярах легких, когда уровень O2, рН и СО2 в крови повышает сродство гемоглобина к О2 по ходу легочных капилляров. В условиях тканей организма эти же факторы метаболизма понижают сродство гемоглобина к О2 и способствуют переходу оксигемоглобина в его восстановленную форму — дезоксигемоглобин. В результате О2 по концентрационному градиенту поступает из крови тканевых капилляров в ткани организма.
Оксид углерода (II) — СО, способен соединяться с атомом железа гемоглобина, изменяя его свойства и реакцию с О2. Очень высокое сродство СО к Нb (в 200 раз выше, чем у О2) блокируют один или более атомов железа в молекуле гема, изменяя сродство Нb к О2.
Под кислородной емкостью крови понимают количество Ог, которое связывается кровью до полного насыщения гемоглобина. При содержании гемоглобина в крови 8,7 ммоль*л-1 кислородная емкость крови составляет 0,19 мл О2 в 1 мл крови (температура 0oC и барометрическое давление 760 мм рт.ст., или 101,3 кПа). Величину кислородной емкости крови определяет количество гемоглобина, 1 г которого связывает 1,36—1,34 мл О2. Кровь человека содержит около 700—800 г гемоглобина и может связать таким образом почти 1 л О2. Физически растворенного в 1 мл плазмы крови О2 очень мало (около 0,003 мл), что не может обеспечить кислородный запрос тканей. Растворимость О2 в плазме крови равна 0,225 мл*л-1*кПа-1.
Обмен О2 между кровью капилляров и клетками тканей также осуществляется путем диффузии. Концентрационный градиент О2 между артериальной кровью (100 мм рт.ст., или 13,3 кПа) и тканями (около 40 мм рт.ст., или 5,3 кПа) равен в среднем 60 мм рт.ст. (8,0 кПа). Изменение градиента может быть обусловлено как содержанием О2 в артериальной крови, так и коэффициентом утилизации О2, который составляет в среднем для организма 30— 40%. Коэффициентом утилизации кислорода называется количество О2, отданного при прохождении крови через тканевые капилляры, отнесенное к кислородной емкости крови.
С другой стороны, известно, что при напряжении О2 в артериальной крови капилляров, равном 100 мм рт.ст. (13,3 кПа), на мембранах клеток, находящихся между капиллярами, эта величина не превышает 20 мм рт.ст. (2,7 кПа), а в митохондриях равна в среднем 0,5 мм рт.ст. (0,06 кПа). Поступление СО2 в легких из крови в альвеолы обеспечивается из следующих источников: 1) из СО2, растворенного в плазме крови (5—10%); 2) из гидрокарбонатов (80—90%); 3) из карбаминовых соединений эритроцитов (5—15%), которые способны диссоциировать.
Для СО2 коэффициент растворимости в мембранах аэрогематического барьера больше, чем для О2, и составляет в среднем 0,231 ммоль*л-1 кПа-1 поэтому СО2 диффундирует быстрее, чем O2. Это положение является верным только для диффузии молекулярного СО2. Большая часть СО2 транспортируется в организме в связанном состоянии в виде гидрокарбонатов и карбаминовых соединений, что увеличивает время обмена СО2, затрачиваемое на диссоциацию этих соединений.
В венозной крови, притекающей к капиллярам легких, напряжение СО2 составляет в среднем 46 мм рт.ст. (6,1 кПа), а в альвеолярном воздухе парциальное давление СО2 равно в среднем 40 мм рт.ст. (5,3 кПа), что обеспечивает диффузию СО2 из плазмы крови в альвеолы легких по концентрационному градиенту.
Эндотелий капилляров проницаем только для молекулярного СО2 как полярной молекулы (О — С — О). Из крови в альвеолы диффундирует физически растворенный в плазме крови молекулярный СО2. Кроме того, в альвеолы легких диффундирует СО2, который высвобождается из карбаминовых соединений эритроцитов благодаря реакции окисления гемоглобина в капиллярах легкого, а также из гидрокарбонатов плазмы крови в результате их быстрой диссоциации с помощью фермента карбоангидразы, содержащейся в эритроцитах.
Молекулярный СО2 проходит аэрогематический барьер, а затем поступает в альвеолы.
В норме через 1 с происходит выравнивание концентраций СО2 на альвеолярно-капиллярной мембране, поэтому за половину времени капиллярного кровотока происходит полный обмен СО2 через аэрогематический барьер. Реально равновесие наступает несколько медленнее. Это связано с тем, что перенос СО2, так же как и О2, ограничивается скоростью перфузии капилляров легких.
Диффузия СО2 из тканей в кровь. Обмен СО2 между клетками тканей с кровью тканевых капилляров осуществляется с помощью следующих реакций: 1) обмена С1- и НСО3- через мембрану эритроцита; 2) образования угольной кислоты из гидрокарбонатов; 3) диссоциации угольной кислоты и гидрокарбонатов.
В ходе газообмена СО2 между тканями и кровью содержание НСОз- в эритроците повышается и они начинают диффундировать в кровь. Для поддержания электронейтральности в эритроциты начнут поступать из плазмы дополнительно ионы С1- Наибольшее количество бикарбонатов плазмы крови образуется при участии карбоангидразы эритроцитов.
Карбаминовый комплекс СО2 с гемоглобином образуется в результате реакции СО2 с радикалом NH2 глобина. Эта реакция протекает без участия какого-либо фермента, т. е. она не нуждается в катализе. Реакция СО2 с Нb приводит, во-первых, к высвобождению Н+; во-вторых, в ходе образования карбаминовых комплексов снижается сродство Нb к О2. Эффект сходен с действием низкого рН. Как известно, в тканях низкое рН потенцирует высвобождение О2 из оксигемоглобина при высокой концентрации СО2 (эффект Бора). С другой стороны, связывание О2 гемоглобином снижает сродство его аминогрупп к СО2 (эффект Холдена).
Каждая реакция в настоящее время хорошо изучена. Например, полупериод обмена С1-и НСО3- равен 0,11—0,16 с при 37 oС. В условиях in vitro образование молекулярного СО2 из гидрокарбонатов происходит чрезвычайно медленно и диффузия этого газа занимает около 5 мин, тогда как в капиллярах легкого равновесие наступает через 1 с. Это определяется функцией фермента карбоангидразы угольной кислоты. В функции карбоангидразы выделяют следующие типы реакций:
СО2+Н2Оß> H2СО3 ß> H++НСО3-
Процесс выведения СО2 из крови в альвеолы легкого менее лимитирован, чем оксигенация крови. Это обусловлено тем, что молекулярный СО2 легче проникает через биологические мембраны, чем О2. По этой причине он легко проникает из тканей в кровь. К тому же карбоангидраза способствует образованию гидрокарбоната. Яды, которые ограничивают транспорт О2 (такие как СО, метгемоглобинобразующие субстанции — нитриты, метиленовый синий, ферроцианиды и др.) не действуют на транспорт СО2. Блокаторы карбоангидразы, например диакарб, которые используются нередко в клинической практике или для профилактики горной или высотной болезни, полностью никогда не нарушают образование молекулярного СО2. Наконец, ткани обладают большой буферной емкостью, но не защищены от дефицита О2. По этой причине нарушение транспорта О2 наступает в организме гораздо чаще и быстрее, чем нарушения газообмена СО2. Тем не менее при некоторых заболеваниях высокое содержание СО2 и ацидоз могут быть причиной смерти.
Измерение напряжения О2 и СО2 в артериальной или смешанной венозной крови производят полярографическими методами с использованием очень небольшого количества крови. Количество газов в крови измеряют после их полного извлечения из пробы крови, взятой для анализа.
Такие исследования выполняют с помощью манометрических приборов типа аппарата Ван-Слайка, или гемоалкариметра (необходимо 0,5—2,0 мл крови) или на микроманометре Холандера (необходимо около 50 мкл крови). 67. Кислородная емкость крови. Механизм транспорта кислорода кровью. Анализ кривой диссоциации оксигемоглобина. Лишь небольшая часть O2 (около 2 %), переносимого кровью, растворена в плазме. Основная его часть транспортируется в форме непрочного соединения с гемоглобином, который у позвоночных содержится в эритроцитах. В молекулы этого дыхательного пигмента входят видоспецифичный белок - глобин — и одинаково построенная у всех животных простетическая группа — гем, содержащая двухвалентное железо. Присоединение кислорода к гемоглобину (оксигенация гемоглобина) происходит без изменения валентности железа, т.е. без переноса электронов, характеризующего истинное окисление. Тем не менее гемоглобин, связанный с кислородом, принято называть окисленным (правильнее — оксигемоглобин), а отдавший кислород — восстановленным (правильнее — дезоксигемоглобин). 1 г гемоглобина может связать 1,36 мл газообразного O2 (при атмосферном давлении). Учитывая, к примеру, что в крови человека содержится примерно 15 г % гемоглобина, 100 мл его крови могут переносить около 21 мл O2. Это так называемая кислородная емкость крови. Оксигенация гемоглобина (иначе говоря, процент, на который используется кислородная емкость крови) зависит от парциального давления O2 в среде, с которой контактирует кровь. Такая зависимость описывается кривой диссоциации оксигемоглобина (рис. 9.1.1). Сложная S-образная форма этой кривой объясняется кооперативным эффектом четырех полипептидных цепей гемоглобина, кислородсвязывающие свойства (сродство к O2) которых различны.
Рис. 9.11. Кривые диссоциации оксигемоглобина: А — кривая насыщения гемоглобина кислородом при нормальном содержании СО2; Б — влияние изменения напряжения СО2 на кривую диссоциации оксигемоглобина; 1 — при низком содержании CO2, 2 — норма, 3 — при высоком содержании СО2 Благодаря такой особенности венозная кровь, проходя легочные капилляры (альвеолярное Р O2 приходится на верхнюю часть кривой), оксигенирируется почти полностью, а артериальная кровь в капиллярах тканей (где Р О2 соответствует крутой части кривой) эффективно отдает O2. Отдаче кислорода способствует содержащийся в эритроцитах 2,3-дифосфоглицерат, синтез которого усиливается при гипоксии и интенсификации окислительного процесса в тканях. Кривая диссоциации оксигемоглобина сдвигается вправо при повышении температуры и при увеличении концентрации водородных ионов в среде, которая, в свою очередь, зависит от Р CO2 (эффект Бора). Поэтому создаются условия для более полной отдачи кислорода оксигемоглобином в тканях, особенно там, где выше интенсивность метаболизма, например в работающих мышцах. Однако и в венозной крови большая или меньшая часть (от 40 до 70 %) гемоглобина остается в оксигенированной форме. Так, у человека каждые 100 мл крови отдают тканям 5−6 мл O2 (так называемая артериовенозная разница по кислороду) и, естественно, на ту же величину обогащаются кислородом в легких. Сродство гемоглобина к кислороду измеряется величиной парциального давления кислорода, при которой гемоглобин насыщается на 50 % (Р 50); У человека оно составляет в норме 26,5 мм рт. ст. для артериальной крови. Параметр Р 50 отражает способность дыхательного пигмента связывать кислород. Этот параметр выше для гемоглобина животных, обитающих в бедной кислородом среде, а также для так называемого фатального гемоглобина, который содержится в крови плода, получающего кислород из крови матери через плацентарный барьер.
68. Транспорт углекислоты кровью. Гидрокарбонатная и каpбаминовая формы связи СО2. Роль карбоангидразы в переносе СО2 кровью. Поступление СО2 в легких из крови в альвеолы обеспечивается из следующих источников: 1) из СО2, растворенного в плазме крови (5—10%); 2) из гидрокарбонатов (80—90%); 3) из карбаминовых соединений эритроцитов (5—15%), которые способны диссоциировать.
Для СО2 коэффициент растворимости в мембранах аэрогематического барьера больше, чем для О2, и составляет в среднем 0,231 ммоль*л-1 кПа-1 поэтому СО2 диффундирует быстрее, чем O2. Это положение является верным только для диффузии молекулярного СО2. Большая часть СО2 транспортируется в организме в связанном состоянии в виде гидрокарбонатов и карбаминовых соединений, что увеличивает время обмена СО2, затрачиваемое на диссоциацию этих соединений.
В венозной крови, притекающей к капиллярам легких, напряжение СО2 составляет в среднем 46 мм рт.ст. (6,1 кПа), а в альвеолярном воздухе парциальное давление СО2 равно в среднем 40 мм рт.ст. (5,3 кПа), что обеспечивает диффузию СО2 из плазмы крови в альвеолы легких по концентрационному градиенту.
Эндотелий капилляров проницаем только для молекулярного СО2 как полярной молекулы (О — С — О). Из крови в альвеолы диффундирует физически растворенный в плазме крови молекулярный СО2. Кроме того, в альвеолы легких диффундирует СО2, который высвобождается из карбаминовых соединений эритроцитов благодаря реакции окисления гемоглобина в капиллярах легкого, а также из гидрокарбонатов плазмы крови в результате их быстрой диссоциации с помощью фермента карбоангидразы, содержащейся в эритроцитах.
Молекулярный СО2 проходит аэрогематический барьер, а затем поступает в альвеолы.
В норме через 1 с происходит выравнивание концентраций СО2 на альвеолярно-капиллярной мембране, поэтому за половину времени капиллярного кровотока происходит полный обмен СО2 через аэрогематический барьер. Реально равновесие наступает несколько медленнее. Это связано с тем, что перенос СО2, так же как и О2, ограничивается скоростью перфузии капилляров легких.
Диффузия СО2 из тканей в кровь. Обмен СО2 между клетками тканей с кровью тканевых капилляров осуществляется с помощью следующих реакций: 1) обмена С1- и НСО3- через мембрану эритроцита; 2) образования угольной кислоты из гидрокарбонатов; 3) диссоциации угольной кислоты и гидрокарбонатов.
В ходе газообмена СО2 между тканями и кровью содержание НСОз- в эритроците повышается и они начинают диффундировать в кровь. Для поддержания электронейтральности в эритроциты начнут поступать из плазмы дополнительно ионы С1- Наибольшее количество бикарбонатов плазмы крови образуется при участии карбоангидразы эритроцитов.
Карбаминовый комплекс СО2 с гемоглобином образуется в результате реакции СО2 с радикалом NH2 глобина. Эта реакция протекает без участия какого-либо фермента, т. е. она не нуждается в катализе. Реакция СО2 с Нb приводит, во-первых, к высвобождению Н+; во-вторых, в ходе образования карбаминовых комплексов снижается сродство Нb к О2. Эффект сходен с действием низкого рН. Как известно, в тканях низкое рН потенцирует высвобождение О2 из оксигемоглобина при высокой концентрации СО2 (эффект Бора). С другой стороны, связывание О2 гемоглобином снижает сродство его аминогрупп к СО2 (эффект Холдена).
Каждая реакция в настоящее время хорошо изучена. Например, полупериод обмена С1-и НСО3- равен 0,11—0,16 с при 37 oС. В условиях in vitro образование молекулярного СО2 из гидрокарбонатов происходит чрезвычайно медленно и диффузия этого газа занимает около 5 мин, тогда как в капиллярах легкого равновесие наступает через 1 с. Это определяется функцией фермента карбоангидразы угольной кислоты. В функции карбоангидразы выделяют следующие типы реакций:
СО2+Н2Оß> H2СО3 ß> H++НСО3-
Процесс выведения СО2 из крови в альвеолы легкого менее лимитирован, чем оксигенация крови. Это обусловлено тем, что молекулярный СО2 легче проникает через биологические мембраны, чем О2. По этой причине он легко проникает из тканей в кровь. К тому же карбоангидраза способствует образованию гидрокарбоната. Яды, которые ограничивают транспорт О2 (такие как СО, метгемоглобинобразующие субстанции — нитриты, метиленовый синий, ферроцианиды и др.) не действуют на транспорт СО2.Блокаторы карбоангидразы, например диакарб, которые используются нередко в клинической практике или для профилактики горной или высотной болезни, полностью никогда не нарушают образование молекулярного СО2. Наконец, ткани обладают большой буферной емкостью, но не защищены от дефицита О2. По этой причине нарушение транспорта О2 наступает в организме гораздо чаще и быстрее, чем нарушения газообмена СО2. Тем не менее при некоторых заболеваниях высокое содержание СО2 и ацидоз могут быть причиной смерти.
Измерение напряжения О2 и СО2 в артериальной или смешанной венозной крови производят полярографическими методами с использованием очень небольшого количества крови. Количество газов в крови измеряют после их полного извлечения из пробы крови, взятой для анализа.
Такие исследования выполняют с помощью манометрических приборов типа аппарата Ван-Слайка, или гемоалкариметра (необходимо 0,5—2,0 мл крови) или на микроманометре Холандера (необходимо около 50 мкл крови). Дыхательный центр и его отделы (дорсальная и вентральная группы респираторных нейронов, пневмотаксический центр). Регуляция дыхания при изменении газового состава крови (с хеморецепторов рефлексогенных зон), при раздражении механорецепторов легких и верхних дыхательных путей Дыхательный центр
Под дыхательным центром следует понимать совокупность нейронов специфических (дыхательных) ядер продолговатого мозга, способных генерировать дыхательный ритм.
В нормальных (физиологических) условиях дыхательный центр получает афферентные сигналы от периферических и центральных хеморецепторов, сигнализирующих соответственно о парциальном давлении О2 в крови и концентрации Н+ во внеклеточной жидкости мозга. В период бодрствования деятельность дыхательного центра регулируется дополнительными сигналами, исходящими из различных структур ЦНС. У человека это, например, структуры, обеспечивающие речь. Речь (пение) может в значительной степени отклонить от нормального уровень газов крови, даже снизить реакцию дыхательного центра на гипоксию или гиперкапнию. Афферентные сигналы от хеморецепторов тесно взаимодействуют с другими афферентными стимулами дыхательного центра, но, в конечном счете, химический, или гуморальный, контроль дыхания всегда доминирует над нейрогенным. Например, человек произвольно не может бесконечно долго задерживать дыхание из-за нарастающих во время остановки дыхания гипоксии и гиперкапнии.
Дыхательный центр выполняет две основные функции в системе дыхания: моторную, или двигательную, которая проявляется в виде сокращения дыхательных мышц, и гомеостатическую, связанную с изменением характера дыхания при сдвигах содержания О2 и СО2 во внутренней среде организма.
Двигательная функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна. Под генерацией дыхательного ритма понимают генерацию дыхательным центром вдоха и его прекращение (переход в экспирацию). Под паттерном дыхания следует понимать длительность вдоха и выдоха, величину дыхательного объема, минутного объема дыхания. Моторная функция дыхательного центра адаптирует дыхание к метаболическим потребностям организма, приспосабливает дыхание в поведенческих реакциях (поза, бег и др.), а также осуществляет интеграцию дыхания с другими функциями ЦНС.
Гомеостатическая функция дыхательного центра поддерживает нормальные величины дыхательных газов (O2, CO2) и рН в крови и внеклеточной жидкости мозга, регулирует дыхание при изменении температуры тела, адаптирует дыхательную функцию к условиям измененной газовой среды, например при пониженном и повышенном барометрическом давлении.
Локализация и функциональные свойства дыхательных нейронов. Нейроны дыхательного центра локализованы в дорсомедиальной и вентролатеральной областях продолговатого мозга и образуют так называемые дорсальную и вентральную дыхательную группу.
Дыхательные нейроны, активность которых вызывает инспирацию или экспирацию, называются соответственно инспираторными и экспираторными нейронами. Инспираторные и экспираторные нейроны иннервируют дыхательные мышцы. В дорсальной и вентральной дыхательной группах продолговатого мозга обнаружены следующие основные типы дыхательных нейронов: 1) ранние инспираторные, которые разряжаются с максимальной частотой в начале фазы вдоха; 2) поздние инспираторные, максимальная частота разрядов которых приходится на конец инспирации; 3) полные инспираторные с постоянной или с постепенно нарастающей активностью в течение фазы вдоха; 4) постинспираторные, которые имеют максимальный разряд в начале фазы выдоха; 5) экспираторные с постоянной или постепенно нарастающей активностью, которую они проявляют во вторую часть фазы выдоха; 6) преинспираторные, которые имеют максимальный пик активности непосредственно перед началом вдоха. Тип нейронов определяется по проявлению его активности относительно фазы вдоха и выдоха.
Нейроны дыхательного центра, активность которых совпадает с ритмом дыхания, но они не иннервируют дыхательные мышцы, называются респираторно-связанными нейронами. К респираторно-связанным нейронам относят клетки дыхательного центра, иннервирующие мышцы верхних дыхательных путей, например гортани.
Дорсальная дыхательная группа (ДДГ) включает в себя симметричные области продолговатого мозга, расположенные вентролатеральнее ядра одиночного пучка (рис. 8.10). Дыхательные нейроны этой группы относятся только к инспираторному типу нейронов и представлены поздними и полными инспираторными нейронами.
Нейроны ДДГ получают афферентные сигналы от легочных рецепторов растяжения по волокнам блуждающего нерва, нейроны которого имеют обширные синаптические связи с другими отделами дыхательного центра и с различными отделами ЦНС. Только часть инспираторных нейронов ДДГ связана аксонами с дыхательными мотонейронами спинного мозга, преимущественно с контралатеральной стороны.
Вентральная дыхательная группа (ВДГ) расположена латеральнее обоюдного ядра продолговатого мозга, или ядра блуждающего нерва. ВДГ подразделяется на ростральную и каудальную части относительно уровня задвижки (obex) продолговатого мозга (см. рис. 8.10).
Ростральная часть ВДГ состоит из инспираторных нейронов разных типов: ранних, полных, поздних инспираторных и постинспираторных. Ранние инспираторные и постинспираторные нейроны ВДГ называются проприобульбарными нейронами, так как они не направляют свои аксоны за пределы дыхательного центра продолговатого мозга и контактируют только с другими типами дыхательных нейронов. Часть полных и поздних инспираторных нейронов направляют свои аксоны к дыхательным мотонейронам спинного мозга, а следовательно, управляют мышцами вдоха.
Каудальная часть ВДГ состоит только из экспираторных нейронов. Все экспираторные нейроны направляют аксоны в спинной мозг. При этом 40% экспираторных нейронов иннервирует внутренние межреберные мышцы, а 60% — мышцы брюшной стенки.
Ростральнее ВДГ локализованы компактной группой экспираторные нейроны (комплекс Бетцингера), аксоны которых связаны только с другими типами нейронов дыхательного центра. Предполагают, что именно эти нейроны синхронизируют деятельность правой и левой половин дыхательного центра.
В непосредственной близости от нейронов ВДГ расположены различные типы респираторно-связанных нейронов, которые иннервируют мышцы верхних дыхательных путей и гортани.
Нейроны дыхательного центра в зависимости от проекции их аксонов подразделяют на три группы: 1) нейроны, иннервирующие мышцы верхних дыхательных путей и регулирующие поток воздуха в дыхательных путях; 2) нейроны, которые синаптически связаны с дыхательными мотонейронами спинного мозга и управляют таким образом мышцами вдоха и выдоха; 3) проприобульбарные нейроны, которые связаны с другими нейронами дыхательного центра и участвуют только в генерации дыхательного ритма.
Другие области локализации дыхательных нейронов. В мосту находятся два ядра дыхательных нейронов: медиальное парабрахиальное ядро и ядро Шатра (ядро Келликера). Иногда эти ядра называют пневмотаксическим центром. В первом ядре находятся преимущественно инспираторные, экспираторные, а также фазавопереходные нейроны, а во втором — инспираторные нейроны. У наркотизированных животных разрушение этих ядер вызывает уменьшение частоты и увеличение амплитуды дыхательных движений. Предполагают, что дыхательные нейроны моста участвуют в механизме смены фаз дыхания и регулируют величину дыхательного объема. В сочетании с двусторонней перерезкой блуждающих нервов разрушение указанных ядер вызывает остановку дыхания на вдохе, или инспираторный апнейзис. Инспираторный апнейзис прерывается редкими, кратковременными и быстрыми выдохами. После выхода животных из наркоза апнейзис исчезает и восстанавливается ритмичное дыхание.
Диафрагмальные мотонейроны. Образуют диафрагмальный нерв. Нейроны расположены узким столбом в медиальной части вентральных рогов от СIII до CV. Диафрагмальный нерв состоит из 700—800 миелинизированных и более 1500 немиелинизированных волокон. Подавляющее количество волокон является аксонами α-мотонейронов, а меньшая часть представлена афферентными волокнами мышечных и сухожильных веретен, локализованных в диафрагме, а также рецепторов плевры, брюшины и свободных нервных окончаний самой диафрагмы.
Мотонейроны сегментов спинного мозга, иннервирующие дыхательные мышцы. На уровне CI—СII вблизи латерального края промежуточной зоны серого вещества находятся инспираторные нейроны, которые участвуют в регуляции активности межреберных и диафрагмальных мотонейронов (см. рис. 8.10).
Мотонейроны, иннервирующие межреберные мышцы, локализованы в сером веществе передних рогов на уровне от TIV до ТX. Причем одни нейроны регулируют преимущественно дыхательную, а другие — преимущественно позно-тоническую активность межреберных мышц. Мотонейроны, иннервирующие мышцы брюшной стенки, локализованы в пределах вентральных рогов спинного мозга на уровне TIV—LIII.
Генерация дыхательного ритма. Спонтанная активность нейронов дыхательного центра начинает появляться к концу периода внутриутробного развития. Об этом судят по периодически возникающим ритмическим сокращениям мышц вдоха у плода. В настоящее время доказано, что возбуждение дыхательного центра у плода появляется благодаря пейсмекерным свойствам сети дыхательных нейронов продолговатого мозга. Иными словами, первоначально дыхательные нейроны способны самовозбуждаться. Этот же механизм поддерживает вентиляцию легких у новорожденных в первые дни после рождения. С момента рождения по мере формирования синаптических связей дыхательного центра с различными отделами ЦНС пейсмекерный механизм дыхательной активности быстро теряет свое физиологическое значение. У взрослых ритм активности в нейронах дыхательного центра возникает и изменяется только под влиянием различных синаптических воздействий на дыхательные нейроны.
Дыхательный цикл подразделяют на фазу вдоха и фазу выдоха относительно движения воздуха из атмосферы в сторону альвеол (вдох) и обратно (выдох). Двум фазам внешнего дыхания соответствуют три фазы активности нейронов дыхательного центра продолговатого мозга: инспираторная, которая соответствует вдоху; постинспираторная, которая соответствует первой половине выдоха и называется пассивной контролируемой экспирацией; экспираторная, которая соответствует второй половине фазы выдоха и называется фазой активной экспирации
Генерация дыхательного ритма происходит в сети нейронов продолговатого мозга, сформированной шестью типами дыхательных нейронов (см. рис. 8.9). Доказано, что сеть основных типов дыхательных нейронов продолговатого мозга способна генерировать дыхательный ритм in vitro в срезах продолговатого мозга толщиной всего 500 мкм, помещенных в искусственную питательную среду.
Инспираторная активность дыхательного центра начинается с мощного стартового разряда ранних инспираторных нейронов, который появляется спонтанно за 100—200 мс до разряда в диафрагмальном нерве. В этот момент ранние инспираторные нейроны полностью освобождаются от сильного торможения со стороны постинспираторных нейронов. Полное растормаживание ранних инспираторных нейронов происходит в момент, когда активируются преинспираторные нейроны дыхательного центра, которые окончательно блокируют разряд экспираторных нейронов.
Стартовый разряд ранних инспираторных нейронов начинает активировать полные инспираторные нейроны, которые способны совозбуждать друг друга. Полные инспираторные нейроны, благодаря этому свойству, поддерживают и увеличивают частоту генерации потенциалов действия в течение фазы вдоха. Именно этот тип дыхательных нейронов создает нарастающую активность в диафрагмальном и межреберных нервах, вызывая увеличение силы сокращения диафрагмы и наружных межреберных мышц.
Ранние инспираторные нейроны в силу особых физиологических свойств их мембраны прекращают генерировать потенциалы действия к середине фазы вдоха. Это моносинаптически растормаживает поздние инспираторные нейроны, поэтому их активность появляется в конце вдоха.
Поздние инспираторные нейроны способны дополнительно активировать в конце вдоха сокращение диафрагмы и наружных межреберных мышц. Одновременно поздние инспираторные нейроны выполняют функцию начального выключения инспирации. В период своей активности они получают возбуждающие стимулы от легочных рецепторов растяжения, которые измеряют степень растяжения дыхательных путей во время вдоха. Максимальный по частоте разряд поздних инспираторных нейронов приходится на момент прекращения активности других типов инспираторных нейронов дыхательного центра.
Прекращение активности всех типов инспираторных нейронов дыхательного центра растормаживает постинспираторные нейроны. Причем процесс растормаживания постинспираторных нейронов начинается гораздо раньше, а именно в период убывания разрядов ранних инспираторных нейронов. С момента появления активности постинспираторных нейронов выключается инспирация и начинается фаза пассивной контролируемой экспирации. Постинспираторные нейроны регулируют степень расслабления диафрагмы в первую половину фазы выдоха. В эту фазу заторможены все другие типы нейронов дыхательного центра. Однако в постинспираторную фазу сохраняется активность респираторно-связанных нейронов дыхательного центра, которые регулируют тонус мышц верхних дыхательных путей, прежде всего гортани.
Вторая половина фазы выдоха, или фаза активной экспирации, полностью зависит от механизма ритмогенеза инспираторнои и пост-инспираторной активности. Например, при быстрых дыхательных движениях постинспираторная фаза может непосредственно переходить в фазу следующей инспирации.
Активность дыхательных мышц в течение трех фаз нейронной активности дыхательного центра изменяется следующим образом (см. рис. 8.11). В инспирацию мышечные волокна диафрагмы и наружных межреберных мышц постепенно увеличивают силу сокращения. В этот же период активируются мышцы гортани, которые расширяют голосовую щель, что снижает сопротивление воздушному потоку на вдохе. Работа инспираторных мышц во время вдоха создает достаточный запас энергии, которая высвобождается в постинспираторную фазу, или в фазу пассивной контролируемой экспирации. В постинспираторную фазу дыхания объем выдыхаемого из легких воздуха контролируется медленным расслаблением диафрагмы и одновременным сокращением мышц гортани. Сужение голосовой щели в постинспираторную фазу увеличивает сопротивление воздушному потоку на выдохе. Это является очень важным физиологическим механизмом, который препятствует спадению воздухоносных путей легких при резком увеличении скорости воздушного потока на выдохе, например при форсированном дыхании или защитных рефлексах кашля и чиханья.
Во вторую фазу выдоха, или фазу активной экспирации, экспираторный поток воздуха усиливается за счет сокращения внутренних межреберных мышц и мышц брюшной стенки. В эту фазу отсутствует электрическая активность диафрагмы и наружных межреберных мышц.
Координация деятельности правой и левой половин дыхательного центра является еще одной функцией дыхательных нейронов. Дыхательный центр имеет дорсальную и вентральную группу нейронов как в правой, так и в левой половине продолговатого мозга и таким образом состоит из двух симметричных половин. Эта функция выполняется за счет синаптического взаимодействия различных типов дыхательных нейронов. Дыхательные нейроны взаимосвязаны как в пределах одной половины дыхательного центра, так и с нейронами противоположной стороны. При этом наибольшее значение в синхронизации деятельности правой и левой половин дыхательного центра имеют проприобульбарные дыхательные нейроны и экспираторные нейроны комплекса Бетцингера. Рефлекторная регуляция дыхания осуществляется благодаря тому, что нейроны дыхательного центра имеют связи с многочисленными механорецепторами дыхательных путей и альвеол легких и рецепторов сосудистых рефлексогенных зон. В легких человека находятся следующие типы механорецепторов: 1) ирритантные, или быстроадаптирующиеся, рецепторы слизистой оболочки дыхательных путей; 2) рецепторы растяжения гладких мышц дыхательных путей; 3) J-рецепторы.
Рефлексы со слизистой оболочки полости носа. Раздражение ирритантных рецепторов слизистой оболочки полости носа, например табачным дымом, инертными частицами пыли, газообразными веществами, водой вызывает сужение бронхов, голосовой щели, брадикардию, снижение сердечного выброса, сужение просвета сосудов кожи и мышц. Защитный рефлекс проявляется у новорожденных при кратковременном погружении в воду. У них возникает остановка дыхания, препятствующая проникновению воды в верхние дыхательные пути.
Рефлексы с глотки. Механическое раздражение рецепторов слизистой оболочки задней части полости носа вызывает сильнейшее сокращение диафрагмы, наружных межреберных мышц, а следовательно, вдох, который открывает дыхательный путь через носовые ходы (аспирационный рефлекс). Этот рефлекс выражен у новорожденных.
Рефлексы с гортани и трахеи. Многочисленные нервные окончания расположены между эпителиальными клетками слизистой оболочки гортани и главных бронхов. Эти рецепторы раздражаются вдыхаемыми частицами, раздражающими газами, бронхиальным секретом, инородными телами. Все это вызывает кашлевой рефлекс, проявляющийся в резком выдохе на фоне сужения гортани и сокращение гладких мышц бронхов, которое сохраняется долгое время после рефлекса.
Кашлевой рефлекс является основным легочным рефлексом блуждающего нерва.
Рефлексы с рецепторов бронхиол. Многочисленные миелинизированные рецепторы находятся в эпителии внутрилегочных бронхов и бронхиол. Раздражение этих рецепторов вызывает гиперпноэ, бронхоконстрикцию, сокращение гортани, гиперсекрецию слизи, но никогда не сопровождается кашлем. Рецепторы наиболее чувствительны к трем типам раздражителей: 1) табачному дыму, многочисленным инертным и раздражающим химическим веществам; 2) повреждению и механическому растяжению дыхательных путей при глубоком дыхании, а также пневмотораксе, ателектазах, действии бронхоконстрикторов; 3) легочной эмболии, легочной капиллярной гипертензии и к легочным анафилактическим феноменам.
Рефлексы с J-рецепторов. В альвеолярных перегородках в контакте с капиллярами находятся особые J-рецепторы. Эти рецепторы особенно чувствительны к интерстициальному отеку, легочной венозной гипертензии, микроэмболии, раздражающим газам и ингаляционным наркотическим веществам, фенилдигуаниду (при внутривенном введении этого вещества). Стимуляция J-рецепторов вызывает вначале апноэ, затем поверхностное тахипноэ, гипотензию и брадикардию.
Рефлекс Геринга — Брейера. Раздувание легких у наркотизированного животного рефлекторно тормозит вдох и вызывает выдох. Перерезка блуждающих нервов устраняет рефлекс. Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких, которые иннервируются миелинизированными волокнами блуждающего нерва.
Рефлекс Геринга — Брейера контролирует глубину и частоту дыхания. У человека он имеет физиологическое значение при дыхательных объемах свыше 1 л (например, при физической нагрузке). У бодрствующего взрослого человека кратковременная двусторонняя блокада блуждающих нервов с помощью местной анестезии не влияет ни на глубину, ни на частоту дыхания.
У новорожденных рефлекс Геринга — Брейера четко проявляется только в первые 3—4 дня после рождения.
Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору больших полушарий и являются единственным источником информации о движениях грудной клетки и дыхательных объемах.
Межреберные мышцы, в меньшей степени диафрагма, содержат большое количество мышечных веретен. Активность этих рецепторов проявляется при пассивном растяжении мышц, изометрическом сокращении и изолированном сокращении интрафузальных мышечных волокон. Рецепторы посылают сигналы в соответствующие сегменты спинного мозга. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которые через γ-мотонейроны повышают активность α-мотонейронов и дозируют таким образом мышечное усилие.
Хеморефлексы дыхания. Рo2 и Рсо2 в артериальной крови человека и животных поддерживается на достаточно стабильном уровне, несмотря на значительные изменения потребления О2 и выделение СО2. Гипоксия и понижение рН крови (ацидоз) вызывают усиление вентиляции (гипервентиляция), а гипероксия и повышение рН крови (алкалоз) — понижение вентиляции (гиповентиляция) или апноэ. Контроль за нормальным содержанием во внутренней среде организма О2, СО2 и рН осуществляется периферическими и центральными хеморецепторами.
Адекватным раздражителем для периферических хеморецепторов является уменьшение Ро2 артериальной крови, в меньшей степени увеличение Рco2 и рН, а для центральных хеморецепторов — увеличение концентрации Н+ во внеклеточной жидкости мозга.
Артериальные (периферические) хеморецепторы. Периферические хеморецепторы находятся в каротидных и аортальных тельцах. Сигналы от артериальных хеморецепторов по синокаротидным и аортальным нервам первоначально поступают к нейронам ядра одиночного пучка продолговатого мозга, а затем переключаются на нейроны дыхательного центра. Ответ периферических хеморецепторов на понижение Рао2 является очень быстрым, но нелинейным. При Рао2 в пределах 80—60 мм рт.ст. (10,6—8,0 кПа) наблюдается слабое усиление вентиляции, а при Раo2 ниже 50 мм рт.ст. (6,7 кПа) возникает выраженная гипервентиляция.
Рaсо2 и pН крови только потенцируют эффект гипоксии на артериальные хеморецепторы и не являются адекватными раздражителями для этого типа хеморецепторов дыхания.
Реакция артериальных хеморецепторов и дыхания на гипоксию. Недостаток О2 в артериальной крови является основным раздражителем периферических хеморецепторов. Импульсная активность в афферентных волокнах синокаротидного нерва прекращается при Рао2 выше 400 мм рт.ст. (53,2 кПа). При нормоксии частота разрядов синокаротидного нерва составляет 10% от их максимальной реакции, которая наблюдается при Раo2 около 50 мм рт.ст. и ниже. Гипоксическая реакция дыхания практически отсутствует у коренных жителей высокогорья и исчезает примерно через 5 лет у жителей равнин после начала их апаптации к высокогорью (3500 м и выше).
Центральные хеморецепторы. Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что такие хеморецепторы находятся в ростральных отделах продолговатого мозга вблизи его вентральной поверхности, а также в различных зонах дорсального дыхательного ядра.
Наличие центральных хеморецепторов доказывается достаточно просто: после перерезки синокаротидных и аортальных нервов у подопытных животных исчезает чувствительность дыхательного центра к гипоксии, но полностью сохраняется реакция дыхания на гиперкапнию и ацидоз. Перерезка ствола мозга непосредственно выше продолговатого мозга не влияет на характер этой реакции.
Адекватным раздражителем для центральных хеморецепторов является изменение концентрации Н* во внеклеточной жидкости мозга. Функцию регулятора пороговых сдвигов рН в области центральных хеморецепторов выполняют структуры гематоэнцефалического барьера, который отделяет кровь от внеклеточной жидкости мозга. Через этот барьер осуществляется транспорт О2, СО2 и Н+ между кровью и внеклеточной жидкостью мозга. Транспорт СО2 и Н+ из внутренней среды мозга в плазму крови через структуры гематоэнцефалического барьера регулируется с участием фермента карбоангидразы.
Реакция дыхания на СО2. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы.
Для определения чувствительности центральных хеморецепторов к изменению рН внеклеточной жидкости мозга используют метод возвратного дыхания. Испытуемый дышит из замкнутой емкости, заполненной предварительно чистым О2. При дыхании в замкнутой системе выдыхаемый СО2 вызывает линейное увеличение концентрации СО2 и одновременно повышает концентрацию Н+ в крови, а также во внеклеточной жидкости мозга. Тест проводят в течение 4—5 мин под контролем содержания СО2 в выдыхаемом воздухе.
Дата добавления: 2015-12-15 | Просмотры: 949 | Нарушение авторских прав |