|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
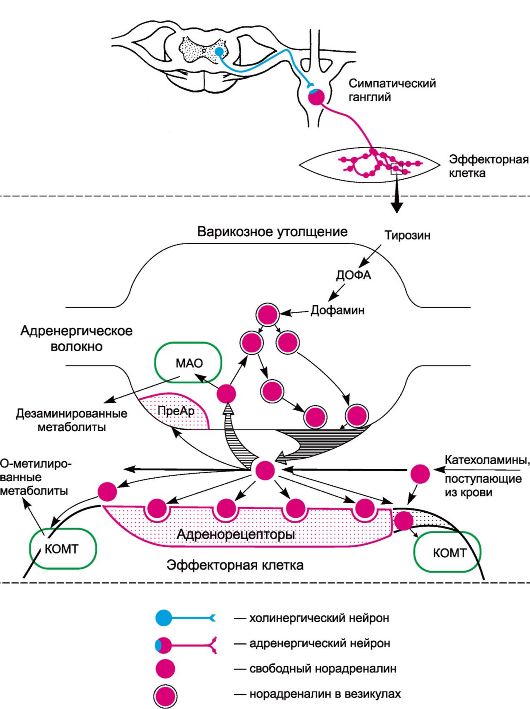
ГЛАВА 4 СРЕДСТВА, ВЛИЯЮЩИЕ НА АДРЕНЕРГИЧЕСКИЕ СИНАПСЫВ адренергических синапсах передача возбуждения осуществляется посредством норадреналина. В пределах периферической иннервации норадреналин принимает участие в передаче импульсов с адренергических волокон на эффекторные клетки (см. рис. 3.1). Адренергические аксоны, подходя к эффектору, разветвляются на тонкую сеть волокон с варикозными утолщениями, выполняющими функцию нервных окончаний. Последние участвуют в образовании синаптических контактов с эффекторными клетками (рис. 4.1). В варикозных утолщениях находятся везикулы (пузырьки), содержащие медиатор норадреналин.
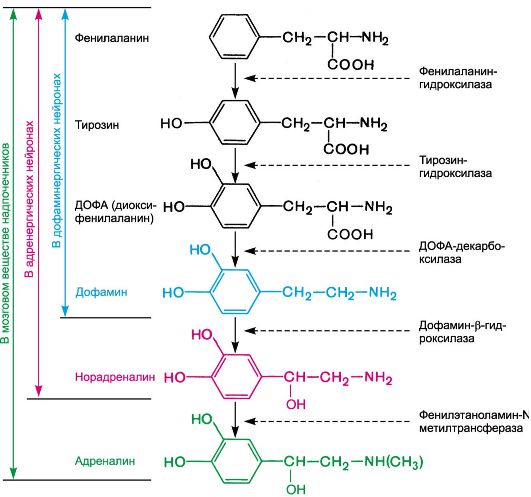
Рис. 4.1. Адренергический синапс (схема). МАО - моноаминоксидаза; КОМТ - катехол-О-метилтрансфераза; ПреАР - пресинаптические адренорецепторы. Биосинтез норадреналина осуществляется в адренергических нейронах из тирозина с участием ряда энзимов. Образование ДОФА и дофамина происходит в цитоплазме нейронов, а норадреналина - в везикулах. Ниже представлены пути биосинтеза дофамина, норадреналина и адреналина (схема 4.1).
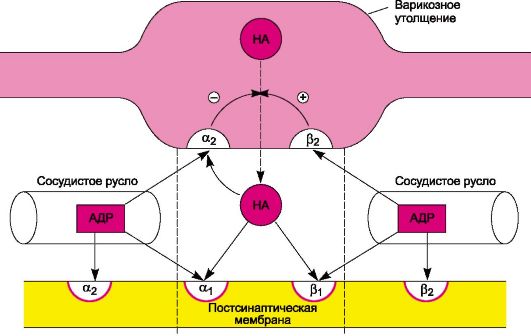
Схема 4.1. Пути биосинтеза дофамина, норадреналина и адреналина. В ответ на нервные импульсы происходит высвобождение норадреналина в синаптическую щель и последующее его взаимодействие с адренорецепторами постсинаптической мембраны. Существующие в организме адренорецепторы обладают неодинаковой чувствительностью к химическим соединениям. Основываясь на этом, выделяют α- и β-адренорецепторы. Основные α-адренорецепторы представлены α 1- и α2-адренорецепторами. α 1-Адренорецепторы локализуются постсинаптически, α2-адренорецепторы - пресинаптически и вне синапсов. Физиологическая роль пресинаптических α2-адренорецепторов заключается в их участии в системе обратной отрицательной связи, регулирующей высвобождение норадреналина. Возбуждение этих рецепторов норадреналином (или другими веществами, обладающими α2-адреномиметической активностью) тормозит высвобождение норадреналина из варикозных утолщений1. α2-Адренорецепторы расположены и на мембранах эффекторных клеток, вне синапсов. Предполагают, что в сосудах они локализуются в неиннервируемом 1 Высказывается предположение, что аналогичную роль играют а2-адренорецепторы, расположенные на окончаниях постганглионарных холинергических волокон (их возбуждение снижает выс- вобождение ацетилхолина). (внутреннем) слое. Очевидно, они возбуждаются в основном циркулирующим в крови адреналином (α1-адренорецепторы активируются преимущественно медиатором норадреналином; рис. 4.2).
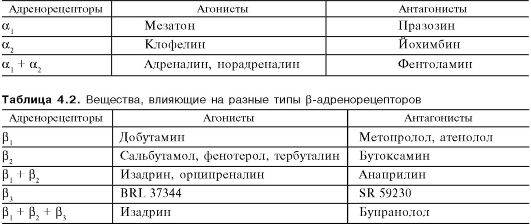
Рис. 4.2. Основная направленность действия норадреналина (НА) и адреналина (АДР) на пресинаптические (α2, β2) и постсинаптические (α1, α2, β1, β2) адренорецепторы. Плюс - стимулирующее действие; минус - угнетающее действие. Среди пост- и внесинаптических β-адренорецепторов выделяют β1-адренорецепторы (например, в сердце), β2-адренорецепторы (в бронхах, сосудах, матке) и β3-адренорецепторы (в жировой ткани). Преимущественная локализация тех или иных β-адренорецепторов приведена для некоторого упрощения материала. Вместе с тем во многих тканях сосуществуют разные типы рецепторов. Так, показано, что в сердце человека и ряда животных наряду с β1-адренорецепторами имеются β2- и β3-адренорецепторы. С другой стороны, в бронхах, помимо β2-адренорецепторов, обнаружены также β1-адренорецепторы. Норадреналин действует преимущественно на иннервируемые β1-адренорецепторы (постсинаптические рецепторы), а адреналин, находящийся в крови, - на β2-адренорецепторы, не имеющие иннервации (внесинаптические рецепторы). Этим объясняется и то, что нейротропные эффекты реализуются в основном через β1-адренорецепторы посредством норадреналина, а гуморальные влияния, например, циркулирующего адреналина, - за счет β2-адренорецепторов. β3-Адренорецепторы активируются катехоламинами в более высоких концентрациях, чем β1- и β2-адренорецепторы. Обна-ружены также пресинаптические β-адренорецепторы (относятся к β2-адренорецепторам). В отличие от аналогичных α-адренорецепторов они осуществляют положительную обратную связь, стимулируя высвобождение норадреналина. Об этом свидетельствует то, что β-агонисты облегчают высвобождение медиатора норадреналина, а β-антагонисты угнетают его. Функционально более важно угнетающее влияние пресинаптических α2-адренорецепторов. Известны вещества, которые избирательно действуют на разные типы адренорецепторов. Это относится как к агонистам, так и к антагонистам (табл. 4.1 и 4.2). Стимуляция определенных постсинаптических адренорецепторов сопровождается типичными для их активации эффектами (табл. 4.3). Так, для стимуляции α-адренорецепторов характерно повышение функции эффекторов (кроме Таблица 4.1. Вещества, влияющие на разные типы α-адренорецепторов
Таблица 4.3. Основные эффекты, связанные со стимуляцией постсинаптических и внесинаптических α- и β-адренорецепторов
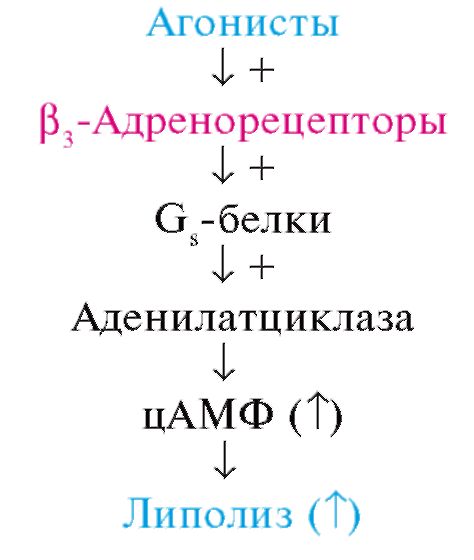
* Активация β3-адренорецепторов снижает силу сокращений желудочков сердца. кишечника, тонус мышц которого падает). При стимуляции β2-адренорецепторов обычно наблюдается снижение функции иннервируемого органа. Возбуждение β1- и β2-адренорецепторов сердца сопровождается увеличением силы и частоты сердечных сокращений, повышением автоматизма и облегчением атриовентрикулярной проводимости. Активация β3-адренорецепторов снижает силу сокращений желудочков. На тромбоцитах имеются а2-адренорецепторы, стимуляция которых повышает агрегацию, и β2-адренорецепторы, выполняющие противоположную функцию (их возбуждение повышает содержание цАМФ). Адренорецепторы принимают участие в регуляции углеводного и жирового обмена. Возбуждение их адреномиметиками сопровождается активацией аденилатциклазы, что приводит к распаду гликогена, высвобождению из жировых тканей свободных жирных кислот. Одна из важных локализаций недавно открытых β3-адренорецепторов - адипоциты жировой ткани. Агонисты этого подтипа рецепторов стимулируют липолиз и термогенез в жировой ткани. Действуют они следующим образом1: 1 (+) - стимулирующее действие; (Τ) - повышение.
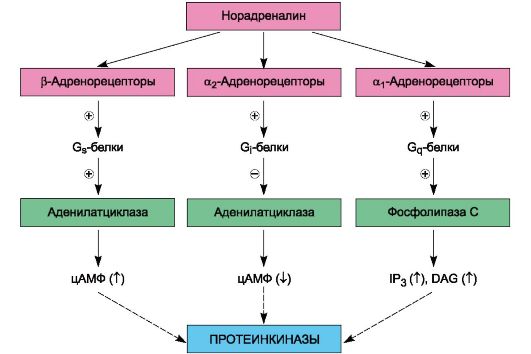
β3-Адренорецепторы иннервируются адренергическими волокнами. К норадреналину они более чувствительны, чем к адреналину. β3-Адренорецепторы обнаружены также в сердце и сосудах, в гладких мышцах пищеварительного тракта, желчного пузыря, в предстательной железе, скелетных мышцах. Их физиологическая роль изучена недостаточно. Агонисты β3-адренорецепторов, по-видимому, перспективны для лечения ожирения, а также в комплексной терапии сахарного диабета. Первые препараты этой группы проходят клинические испытания. Количественное соотношение в тканях а- и β-адренорецепторов различно. Так, в сосудах кожи, почек и кишечника, сфинктерах желудочно-кишечного тракта, трабекулах селезенки преобладают α-адренорецепторы. В сердце, мышцах бронхов, сосудах скелетных мышц в основном находятся β-адренорецепторы. Локализацией и соотношением а- и β-адренорецепторов определяются эффект раздражения адренергических (симпатических) нервов, а также реакция на адреномиметические вещества, возбуждающие а- и β-адренорецепторы. Строение адренорецепторов изучено недостаточно полно. В отношении β1- и а2-адренорецепторов имеются данные, что они посредством G-белков функционально взаимосвязаны с ферментом аденилатциклазой, локализованной в мембране эффекторных клеток и обеспечивающей синтез циклического 3',5'-адено- зинмонофосфата (цАМФ; рис. 4.3; схема 4.2). α1-Адренорецепторы активируют G-белки, связанные с фосфолипазой С. Действие норадреналина на адренорецепторы кратковременно. Это объясняется главным образом быстрым захватом окончаниями адренергических волокон (так называемый нейрональный захват) до 75-80% медиатора, находящегося в синаптической щели, и последующим его депонированием. Катаболизм свободного норадреналина в адренергических окончаниях регулируется ферментом МАО, локализованной в основном в митохондриях и, очевидно, в мембранах везикул. Под влиянием МАО происходит окислительное дезаминирование норадреналина. Метаболизм выделившегося из нервных окончаний норадреналина, а также циркулирующих катехоламинов осуществляется в основном цитоплазматическим ферментом эффекторных клеток - катехол-О-метилтрансферазой (КОМТ). Под влиянием этого фермента происходит О-метилирование катехоламинов. Небольшие количества медиатора подвергаются экстранейрональному захвату эффекторными клетками (гладкими мышцами и др.). При экстранейрональном захвате норадреналин быстро метаболизируется при участии КОМТ и МАО. Таким образом, баланс норадреналина зависит от его синтеза, депонирования, нейронального и экстранейронального захвата, а также от энзиматических превращений.
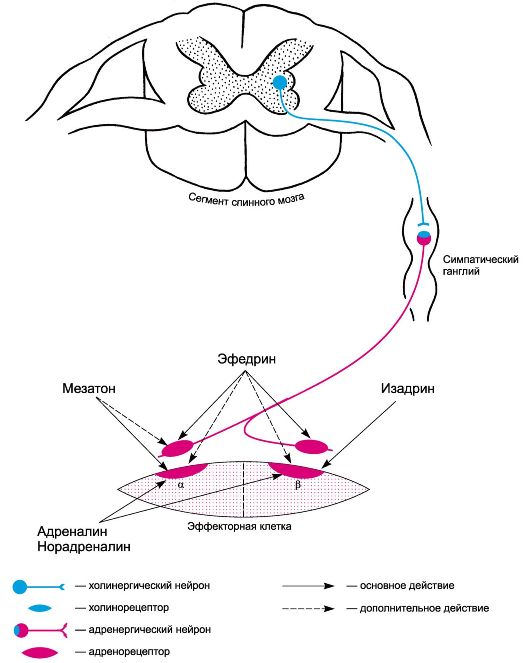
Рис. 4.3. Основная направленность действия адреномиметиков на постсинаптические адренорецепторы. Примечание. Под действием на варикозные утолщения подразумевается симпатомиметическая активность веществ. Возможности фармакологического воздействия на адренергическую передачу нервных импульсов довольно разнообразны. Направленность действия веществ может быть следующей: 1) влияние на синтез норадреналина; 2) нарушение депонирования норадреналина в везикулах и цитоплазме пресинаптических окончаний; 3) угнетение ферментативной инактивации норадреналина; 4) влияние на выделение норадреналина из окончаний; 5) нарушение процесса обратного захвата норадреналина пресинаптическими окончаниями; 6) угнетение экстранейронального захвата норадреналина; 7) непосредственное воздействие на адренорецепторы.
Схема 4.2. Пути сопряжения с эффектором разных подтипов адренорецепторов при влиянии на них норадреналина. IP3 - инозитол 1,4,5-трифосфат, DAG - диацилглицерол, ↓ - снижение концентрации, ↑ - повышение концентрации, о G-белках см. в разделе II.5. Так, синтез норадреналина угнетает α-метил-п-тирозин (тормозит тирозингидроксилазу). Блокируя транспортные системы мембран везикул, резерпин угнетает проникновение в везикулы дофамина и обратный захват норадреналина везикулами. В связи с этим уменьшается содержание депонированного в них норадреналина. Снижение содержания норадреналина в пресинаптических окончаниях отмечается также при применении октадина. Неизбирательным ингибитором МАО является ниаламид, а активность КОМТ угнетают энтакапон, толкапон. Процесс высвобождения норадреналина из пресинаптических окончаний может быть изменен. Одни вещества стимулируют его выделение (например, тирамин, эфедрин), другие уменьшают (октадин, орнид). Нейрональный захват норадреналина адренергическими окончаниями угнетается при введении имизина, кокаина (при этом увеличивается концентрация медиатора в синаптической щели), экстранейрональный захват подавляется метанефрином, феноксибензамином. Наиболее часто в медицинской практике используют вещества, влияющие на адренорецепторы. Вещества, стимулирующие адренорецепторы, называют адреномиметиками, а угнетающие их - адреноблокаторами. С учетом преимущественной локализации действия основные средства, влияющие на передачу возбуждения в адренергических синапсах, подразделяют на следующие группы. 1. Вещества, действующие непосредственно на адренорецепторы: а) адреномиметики прямого действия - норадреналина гидротартрат, адреналина гидрохлорид, изадрин и др.; б) адреноблокаторы - фентоламин, анаприлин и др. 2. Вещества пресинаптического действия, влияющие на высвобождение и(или) депонирование норадреналина: а) симпатомиметики или адреномиметики непрямого действия1 - тирамин, эфедрина гидрохлорид2; б) симпатолитики - октадин, резерпин. Исходя из тропности адреномиметиков и адреноблокаторов в отношении α- и β-адренорецепторов, их можно систематизировать следующим образом. Адреномиметические средства3 • Стимулирующие α - и β-адренорецепторы Адреналина гидрохлорид4 Норадреналина гидротартрат4 (или гидротартрат) (β 1, β 2, α1, α2)(α1, α2, β 1) • Стимулирующие преимущественно α-адренорецепторы Мезатон (α1) Нафтизин (α2) Галазолин (α 2) • Стимулирующие преимущественно β -адренорецепторы Изадрин (β β 2) Сальбутамол (β 2) Добутамин (β 1) Фенотерол (β 2) Тербуталин (β 2) Дата добавления: 2015-12-16 | Просмотры: 1653 | Нарушение авторских прав |