|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ГЛАВА 2. КРАТКИЕ СВЕДЕНИЯ ПО АНАТОМИИ НЕРВНОЙ СИСТЕМЫ2.1. Спинной мозг Спинной мозг (medulla spinalis) расположен в позвоночном канале. На уровне I шейного позвонка и затылочной кости спинной мозг переходит в продолговатый, а книзу тянется до уровня I-II поясничных позвонков (рис. 2.1). По длиннику спинного мозга выделяют шейное и пояснично-крестцовое утолщения. Спинной мозг состоит из 31 - 32 сегментов: 8 шейных (CI - CIV), 12 грудных (ТhI-ТhXII), 5 поясничных (LI-LV), 5 крестцовых (SI-SV) и 1-2 рудиментарных копчиковых (СоI - СоII). Шейное утолщение соответствует сегментам СV-ТhI и обеспечивает иннервацию верхних конечностей. Пояснично-крестцовое утолщение соответствует сегментам LI-SI-II и иннервирует нижние конечности. Ниже поясничного утолщения спинной мозг истончается, образуя мозговой конус, который заканчивается концевой нитью, доходящей до копчиковых позвонков. Сегментом называется участок спинного мозга, которому принадлежит одна пара спинномозговых корешков (передних и задних). Передний корешок спинного мозга содержит двигательные волокна, задний корешок - чувствительные волокна. Соединяясь в области межпозвонкового узла, они образуют смешанный спинномозговой нерв. Спинной мозг несколько короче позвоночного канала. В связи с этим в верхних отделах спинного мозга его корешки идут горизонтально. Затем, начиная с грудного отдела, они несколько спускаются книзу, прежде чем выйти из соответствующих межпозвоночных отверстий. В нижних отделах корешки идут прямо вниз, формируя конский хвост. На поверхности спинного мозга видны передняя срединная щель, задняя срединная борозда, симметрично расположенные передние и задние латеральные борозды. Между передней срединной щелью и передней латеральной бороздой располагается передний канатик (funiculus anterior), между передней и задней латеральными бороздами - боковой канатик (funiculus lateralis), между задней латеральной бороздой и задней срединной бороздой - задний канатик (funiculus posterior), который в шейной части спинного мозга делится неглубокой промежу-
точной бороздой на тонкий пучок (fasciculus gracilis), прилежащий к задней срединной борозде и расположенный кнаружи от него клиновидный пучок (fasciculus cuneatus). Канатики содержат проводящие пути, из передней латеральной борозды выходят передние корешки, в области задней боковой борозды в спинной мозг входят задние корешки. На поперечном срезе (см. рис. 2.2) в спинном мозге выделяются серое вещество, расположенное в центральных отделах спинного мозга, и белое вещество, лежащее на его периферии. Серое вещество на поперечном срезе напоминает по форме бабочку с раскрытыми крыльями или букву «Н». В сером веществе спинного мозга выделяют более массивные, широкие и короткие передние рога и более тонкие, удлиненные задние рога. В грудных отделах выявляется боковой рог, который меньше выражен в поясничном и шейном отделах спинного мозга. Правая и левая половины спинного мозга симметричны и соединены спайками из серого и белого вещества. Кпереди от центрального канала находится передняя серая спайка (comissura grisea anterior), далее передняя белая спайка (comissura alba anterior); кзади от центрального канала последовательно располагаются задняя серая спайка (comissura grisea posterior) и задняя белая спайка (comissura alba posterior). В передних рогах спинного мозга находятся крупные двигательные нейроны, аксоны которых идут в передние корешки и иннервируют поперечнополосатую мускулатуру шеи, туловища и конечностей. Первичные чувствительные клетки располагаются в спинномозговых (межпозвоночных) узлах, представляющих собой утолщения задних корешков. Такая нервная клетка имеет один отросток, который, отходя от нее, делится на две ветви. Одна из них идет на периферию, где получает раздражение от рецеп-
1 - передняя срединная щель; 2 - задний рог: а - верхушка; б - головка; в - шейка; 3 - студенистое вещество; 4 - задний канатик; 5 - задняя срединная борозда; 6 - тонкий пучок; 7 - клиновидный пучок; 8 - задняя срединная перегородка; 9 - боковой канатик; 10 - центральный канал; 11 - передний рог; 12 - передний канатик торов кожи, мышц, сухожилий или внутренних органов, а по другой ветви эти импульсы передаются в спинной мозг. В зависимости от вида раздражения и проводящего пути, по которому оно передается, афферентные волокна, входящие в спинной мозг через задний корешок, могут оканчиваться на клетках задних рогов либо непосредственно проходят в белое вещество спинного мозга. Таким образом, клетки передних рогов осуществляют двигательные функции, клетки задних рогов - чувствительные, в боковых рогах локализуются сегментарные вегетативные клетки. На уровне сегментов СVIII-LIII в боковых рогах располагаются клетки симпатической нервной системы, на уровне сегментов SII-SIV локализован спинальный парасимпатический центр, обеспечивающий регуляцию функции тазовых органов. Белое вещество спинного мозга состоит из волокон восходящих и нисходящих проводящих путей (рис. 2.3), осуществляющих взаимосвязь как различных уровней спинного мозга между собой, так и всех вышележащих отделов ЦНС со спинным мозгом. В передних канатиках спинного мозга расположены проводящие пути, участвующие в осуществлении двигательных функций: 1) передний корково-спинномозговой (пирамидный) путь
(неперекрещенный), идущий в основном от двигательной области коры большого мозга и заканчивающийся на клетках передних рогов; 2) преддверно-спинномозговой (вестибулоспинальный) путь, идущий от латерального вестибулярного ядра той же стороны и заканчивающийся на клетках передних рогов; 3) покрышечно-спинномозговой путь, начинающийся в верхних холмиках четверохолмия противоположной стороны и заканчивающийся на клетках передних рогов; 4) передний ретикулярно-спинномозговой путь, идущий от клеток ретикулярной формации мозгового ствола той же стороны и заканчивающийся на клетках переднего рога. Кроме того, вблизи от серого вещества проходят волокна, осуществляющие связь различных сегментов спинного мозга между собой. В боковых канатиках спинного мозга располагаются как двигательные, так и чувствительные пути. К двигательным путям относятся: 1) латеральный корково-спинномозговой (пирамидный) путь (перекрещенный), идущий от двигательной области коры большого мозга и заканчивающийся на клетках передних рогов противоположной стороны; 2) руброспинальный путь, идущий от красного ядра и заканчивающийся на клетках передних рогов противоположной стороны; 3) ретикулярно-спинномозговые пути, идущие преимущественно от гигантоклеточного ядра ретикулярной формации противоположной стороны и заканчивающиеся на клетках передних рогов; 4) оливоспинномозговой путь, соединяющий нижние оливы с двигательным нейроном переднего рога. К афферентным, восходящим проводникам бокового канатика относятся: 1) задний (дорсальный, неперекрещенный) спинно-мозжечковый путь, идущий от клеток заднего рога и заканчивающийся в коре мозжечка; 2) передний (перекрещенный) спинно-мозжечковый путь, идущий от клеток задних рогов и заканчивающийся в черве мозжечка; 3) латеральный спинно-таламический путь, идущий от клеток задних рогов и заканчивающийся в таламусе. Кроме того, в боковом канатике проходят спинно-покрышечный, спиноретикулярный, спинооливный пути. В задних канатиках спинного мозга располагаются афферентные тонкий и клиновидный пучки. Волокна, входящие в них, начинаются в межпозвонковых узлах и заканчиваются соответственно в ядрах тонкого и клиновидного пучков, располагающихся в нижнем отделе продолговатого мозга. Таким образом, на уровне спинного мозга замыкаются рефлекторные дуги: возбуждение, приходящее по волокнам задних корешков, подвергается анализу и передается на клетки переднего рога для реализации ответа. Рефлекс может осуществляться при наличии трех звеньев: • афферентной части, в которую входят рецепторы и проводящие пути, передающие раздражение в нервные центры; • центральной части рефлекторной дуги, где формируется реакция; • эффекторной части, реализующей реакцию через скелетную мускулатуру, гладкие мышцы и железы. Спинной мозг, таким образом, является одним из первых этапов, на котором осуществляются анализ и синтез раздражений как из внутренних органов, так и от рецепторов кожных покровов, мускулатуры, слизистых оболочек. Наряду с этим спинной мозг передает приходящие афферентные импульсы в вышележащие отделы. Сегментарный вегетативный аппарат спинного мозга участвует в реализации вегетативно-трофических влияний, симпатической иннервации мышц глаза, регуляции деятельности тазовых органов (мочеиспускания и дефекации). Периферические нервы спинного мозга Корешки спинного мозга имеют сегментарное распределение и разделяются на задние (несущие чувствительные волокна) и передние (несущие двигательные волокна). Вблизи от спинного мозга по ходу заднего корешка расположен чувствительный межпозвонковый узел (спинномозговой, или спинальный), в котором находятся первые чувствительные нейроны всех систем поверхностной и глубокой чувствительности. За спинномозговым узлом на уровне каждого сегмента передний и задний корешки сливаются в общий ствол, образуя смешанный спинномозговой нерв (рис. 2.4). Спинномозговые нервы по выходе из межпозвоночного отверстия делятся на ветви: переднюю (иннервирует кожу и мышцы конечностей и передней поверхности туловища), заднюю (иннервирует заднюю поверхность туловища), оболочечную (иннервирует оболочки спинного мозга) и соединительную, направляющуюся к симпатическим узлам (см. рис. 2.4). Передние ветви нескольких соседних сегментов верхнешейного уровня, шейного и пояснично-крестцового утолщений спинного мозга объединяются в сплетения, из которых образуются периферические нервы. Таким обра-
1 - задний рог; 2 - задний канатик; 3 - задняя срединная борозда; 4 - задний корешок; 5 - спинномозговой узел; 6 - ствол спинномозгового нерва; 7 - внутренняя ветвь задней ветви; 8 - наружная ветвь задней ветви; 9 - задняя ветвь; 10 - передняя ветвь; 11 - белые соединительные ветви; 12 - оболочечная ветвь; 13 - серые соединительные ветви; 14 - узел симпатического ствола; 15 - передняя срединная щель; 16 - передний рог; 17 - передний канатик; 18 - передний корешок; 19 - передняя серая спайка; 20 - центральный канал; 21 - боковой канатик; 22 - постганглионарные волокна. Синим цветом обозначены чувствительные волокна, красным - двигательные, зеленым - белые соединительные ветви, фиолетовым - серые соединительные ветви зом, периферические нервы чаще являются смешанными, так как в их составе находятся двигательные, чувствительные и вегетативные волокна. Шейное сплетение (рис. 2.5) образуется из передних ветвей спинномозговых нервов четырех верхних шейных сегментов. Его периферические нервы иннервируют кожу и глубокие мышцы шеи, частично грудинно-ключично-сосцевидную и трапециевидную мышцы, а также диафрагму (сегменты СIII-СV, диафрагмальный нерв). Плечевое сплетение (рис. 2.6) образуется из передних ветвей спинномозговых нервов шейных сегментов СV-СVIII и грудных ThI-ThII. Его периферические нервы осуществляют двигательную и чувствительную иннервацию плечевого пояса и верхних конечностей.
Стволы спинномозговых нервов (корни сплетения) в сегментах CI - CV; X - блуждающий нерв; XI - добавочный нерв; XII - подъязычный нерв: 1 - подбородочноподъязычная мышца; 2 - щитоподъязычная мышца; 3 - грудинно-подъязычная мышца; 4 - грудинно-щитовидная мышца; 5 - лопаточно-подъязычная мышца; 6 - верхний шейный узел симпатического ствола; 7 - верхний корешок шейной петли; 8 - шейная петля; 9 - нижний корешок шейной петли; 10 - диафрагмальный нерв; 11 - ветвь к плечевому сплетению; 12 - медиальные надключичные нервы; 13 - промежуточные надключичные нервы; 14 - латеральные (задние) надключичные нервы; 15 - трапециевидная мышца; 16 - поперечный нерв шеи; 17 - грудинноключично-сосцевидная мышца; 18 - малый затылочный нерв; 19 - большой ушной нерв; 20 - оболочечные веточки подъязычного нерва; 21 - задние двигательные ветви к мышцам спины. Синим цветом обозначены чувствительные волокна, красным - двигательные, зеленым - симпатические, желтым - черепные нервы
I - верхний ствол; II - средний ствол; III - нижний ствол; IV - латеральный пучок; V - задний пучок; VI - медиальный пучок; надключичная часть: 1 - ветвь к диафрагмальной мышце; 2 - дорсальный нерв лопатки (CV); 3 - подключичный нерв (CV, CVI); 4 - надлопаточный нерв (CIV-CVI); 5 - длинный грудной нерв (CV-CVII); подключичная часть: 6 - мышечно-кожный нерв (CIV-CVI); 7 - срединный нерв (CV-CVIII, ThI); 8 - латеральный грудной нерв (CV-CVII); 9 - подмышечный нерв (CV, CVI); 10 - лучевой нерв (CV-CVIII, ThI); 11 - медиальный грудной нерв (CVIII-ThII); 12 - грудоспинной нерв (CVI-CVIII); 13 - подлопаточный нерв (CV, CVI); 14 - локтевой нерв (CVIII, ThI); 15 - медиальный кожный нерв предплечья (CVIII, ThI); 16 - медиальный кожный нерв плеча (ThI); 17 - межреберно-плечевые нервы; 18 - межреберный нерв (II); 19 - межреберный нерв (I); 20 - задние двигательные ветви спинномозговых нервов к передней, средней и задней лестничным мышцам и длинным мышцам шеи. Красным цветом обозначены задние, розовым - передние разветвления трех стволов Передние ветви грудных нервов не формируют сплетения, а образуют обособленные нервы, иннервирующие кожу передней поверхности туловища, межреберные мышцы, мышцы, поднимающие ребра, и мышцы живота. Поясничное сплетение (рис. 2.7) образуется передними ветвями спинномозговых нервов ТhXII-LIV-сегментов, крестцовое сплетение LIV-SIII- сегментов (рис. 2.8). Их периферические нервы осуществляют двигательную и чувствительную иннервацию мускулатуры тазового пояса, нижних конечностей, органов таза. Последний крестцовый и копчиковые спинномозговые нервы (Sv-СоI-II) дают начало нескольким тонким нервам, иннервирующим кожные покровы верхушки копчика. 2.2. Головной мозг Ствол головного мозга Ствол мозга продолжает спинной мозг вверх, в полость черепа, и состоит из продолговатого мозга, моста и среднего мозга (снизу вверх). На сагиттальном срезе в стволе мозга можно выделить основание, покрышку и крышу (рис. 2.9). В основании проходят главным образом нисходящие эфферентные пути. В покрышке расположены ядра черепных нервов, чувствительные (афферентные) пути, ретикулярная формация. Крышей среднего мозга служит пластинка четверохолмия, в определенной степени - мозжечок, онтогенетически связанный со структурами моста. Ядра III и IV черепных нервов располагаются на уровне верхних и нижних бугорков среднего мозга соответственно, ядра V-VIII черепных нервов - на уровне моста, ядра IX-XII - на уровне продолговатого мозга (каудальная группа). Ядра черепных нервов являются аналогами сегментарных структур спинного мозга: двигательные соответствуют передним рогам спинного мозга, чувствительные - задним рогам, вегетативные - боковым рогам (рис. 2.10). Продолговатый мозг Продолговатый мозг (medulla oblongata) является продолжением спинного мозга. Спинной мозг переходит в продолговатый мозг постепенно, без четкой границы. Условной границей перехода спинного мозга в продолговатый является перекрест пирамид (decussatio pyramidum).
1 - ветвь к поясничному сплетению; 2 - пояснично-крестцовый ствол; 3 - верхний ягодичный нерв (LIV, LV, SI); 4 - нижний ягодичный нерв (LV, SI, SII); 5 - задний кожный нерв бедра (SI-SIII); 6 - седалищный нерв; 7 - общий малоберцовый нерв (LIV, LV, SI, Sn); 8 - большеберцовый нерв (LIV, LV, SI-SIII); 9 - мышечные ветви к подколенным мышцам; 10 - мышечные ветви к внутренней запирательной и верхней близнецовой мышцам (LV, SI, SII); 11 - мышечные ветви к квадратной мышце бедра и нижней близнецовой мышце (LIV, LV, SI); 12 - нижние нервы ягодицы; 13 - ветвь к копчиковому сплетению; 14 - задние двигательные ветви спинномозговых нервов к грушевидной мышце (SI, SII обозначены звездочками)
Макроскопически в продолговатом мозге на его вентральной поверхности выделяется передняя срединная щель. Латеральнее от нее четко расположены пирамиды (pyramides), с латеральной стороны ограниченные передними латеральными бороздами. Кнаружи от них располагаются возвышения нижнего оливного ядра. Несколько кзади расположенные задние канатики, разделенные задней срединной бороздой, образуют два отчетливо выраженных возвышения. На этом уровне расположено возвышение, соответствующее расположению ядра тонкого пучка. Клиновидный пучок заканчивается клиновидным ядром (nucl. cuneatus), которое образует небольшое возвышение на дорсальной поверхности продолговатого мозга. Снаружи клиновидный пучок ограничивается задней латеральной бороздой. Из передней боковой борозды выходят корешки подъязычного нерва (n. hypoglossus, XII), из задней боковой борозды выходят каудально волокна языкоглоточного нерва (n. glossopharyngeus, IX), оральнее - блуждающего (n. vagus, X) и добавочного нервов (n. accessorius, XI). Оральнее продолговатый мозг переходит в мост (pons). Граница между ними на базальной поверхности мозга весьма четкая: мостомозжечковые волокна моста образуют мощный пучок, который располагается перпендикулярно длинной оси продолговатого мозга и дорсально образует средние мозжечковые ножки. Пучками мостомозжечковых волокон мост также четко отделяется от среднего мозга, в который он переходит. Дорсально расположенный продолговатый мозг и мост образуют дно Iy желудочка, выстланного эпендимой. Iy желудочек, сужаясь кпереди, переходит в сильвиев водопровод, через который он сообщается с III
1 - парасимпатические ядра глазодвигательного нерва; 2 - ядра глазодвигательного нерва; 3 - ядро бокового нерва; 4 - ядро среднемозгового пути тройничного нерва; 5 - двигательное ядро тройничного нерва; 6 - мостовое ядро тройничного нерва; 7 - ядро отводящего нерва; 8 - ядро лицевого нерва; 9 - вестибулярные ядра; 10 - улитковые ядра; 11- лицевой нерв; 12 - верхнее и нижнее слюноотделительные ядра; 13 - ядро подъязычного нерва; 14 - двойное ядро; 15 - ядро спинномозгового пути тройничного нерва; 16 - ядро одиночного пути; 17 - добавочный нерв; 18 - заднее ядро блуждающего нерва; 19 - ядро добавочного нерва; 20 - задвижка; 21 - задняя срединная борозда; 22 - нежный пучок; 23 - клиновидный пучок; 24 - задняя промежуточная борозда; 25 - нижний угол ромбовидной ямки; 26 - треугольник блуждающего нерва; 27 - треугольник подъязычного нерва; 28 - нижний мозговой парус желудочком; кроме того, IV желудочек имеет 3 отверстия, благодаря которым цереброспинальная жидкость может сообщаться с субарахноидальным пространством: два симметричных отверстия сбоку (отверстия Люшки), в области латеральных карманов, и одно несимметричное отверстие, находящееся в задней части крыши (отверстие Мажанди). Боковые стенки IV желудочка образуются соответственно нижними, средними и верхними мозжечковыми ножками, крыша - нижним и верхним мозговыми парусами. Дорсальнее располагается мозжечок, прикрывающий IV желудочек. Изнутри к нижнему мозговому парусу прилежит сосудистая основа IV желудочка. Дно IV желудочка имеет форму ромба (ромбовидная ямка). Верхние стороны ромба ограничены верхними ножками мозжечка, нижние - нижними ножками. Верхний и нижний углы ромба соединяются продольной срединной бороздой. Из боковых углов ромбовидной ямки к середине идут парные мозговые полоски, разделяющие ее на верхний и нижний треугольники. Условно можно считать, что верхний треугольник соответствует мосту, нижний - продолговатому мозгу. В области ромбовидной ямки находятся ядра V-XII черепных нервов. В ромбовидной ямке от каудального отдела продолговатого мозга тянется крупное ядро подъязычного нерва (XII), осуществляющее двигательную иннервацию языка. Оно располагается несколько латеральнее средней линии и представлено колонкой крупных многоугольных нервных клеток. Латеральнее от него также почти на всем протяжении продолговатого мозга тянется заднее ядро блуждающего нерва (X). Заднее ядро блуждающего нерва состоит из клеток средней величины, веретенообразной или угловатой формы, а также мелких многоугольных клеток. Латеральнее заднего ядра блуждающего нерва располагается ядро одиночного пути (nucl. tractus solitarii), которое тянется от нижнего отдела продолговатого мозга до нижнего отдела моста. В этом ядре заканчиваются афферентные волокна блуждающего и языкоглоточного нервов. В нижнем отделе продолговатого мозга латеральные части его покрышки заняты тонким (nucl. gracilis) и клиновидным (nucl. cuneatus) ядрами, в которых заканчиваются волокна тонкого и клиновидного пучков, идущие из спинного мозга. В наружной части клиновидного пучка на уровне нижнего отдела продолговатого мозга начинает формироваться добавочное клиновидное ядро (nucl. cuneatus accessorius). Волокна от добавочного клиновидного ядра, оливомозжечкового пучка и других более мелких систем начинают формировать нижние мозжечковые ножки. Вентральнее от ядер тонкого и клиновидного пучков располагается ядро спинномозгового пути тройничного нерва (nucl. tractus spinalis n. trigemini), которое тянется через каудальный отдел моста и весь продолговатый мозг. Это ядро окружает постепенно истончающийся пучок волокон (корешок тройничного нерва), который достаточно развит в каудальном отделе моста и верхнем отделе продолговатого мозга и постепенно исчезает в каудальном отделе продолговатого мозга. В клетках ядра спинномозгового пути тройничного нерва осуществляется рецепция болевых ощущений и температуры лица. Средний отдел продолговатого мозга (рис. 2.11) на всех уровнях занимают хорошо выраженные ядра ретикулярной формации. На уровне продолговатого мозга они представлены центральным ретикулярным ядром, которое может быть разделено на вентральное и дорсальное. Характерным образованием для продолговатого мозга является нижнее оливное ядро (nucl. olivaris caudalis), которое осуществляет важную роль в координации движений и имеет тесные связи с мозжечком. К нижнему оливному ядру прилежат заднее и медиальное добавочные оливные ядра (nucl. olivaris accessorius posterior et nucl. olivaris accessorius medialis). Они окружены слоем волокон - капсулой нижней оливы, основную часть которой составляют таламооливные и руброоливные волокна, образующие центральный пучок покрышки.
Кнаружи от нижних оливных ядер в латеральных отделах продолговатого мозга располагаются волокна руброспинального, переднего спинно-мозжечкового, латерального спинноталамического и оливоспинномозгового путей. Между оливными ядрами, дорсальнее пирамид, находятся волокна медиальной петли (lemniscus medialis), которая образована перекрещивающимися волокнами от ядер клиновидного и тонкого пучков. Эти волокна идут через ядра ретикулярной формации и называются внутренними дугообразными волокнами (fibrae arcuatae internae). Над перекрестом внутренних дугообразных волокон, кпереди от ядра подъязычного нерва, лежит медиальный продольный пучок (fasciculus longitudinalis medialis), осуществляющий взаимосвязь многих ядер ствола мозга между собой. В латеральном отделе центрального ретикулярного ядра, на границе между его дорсальным и вентральным подъядрами, располагаются крупные клетки двойного ядра (nucl. ambiguus). Вентральные отделы продолговатого мозга заняты пирамидами, по переднему и медиальному краям которых лежат дугообразные ядра. Около задней латеральной борозды, дорсолатеральнее от нижнего оливного ядра, проходят передний спинно-мозжечковый путь, латеральный спинно-таламический и красноядерно-спинномозговой пути. Задний спинно-мозжечковый путь на этом уровне полностью перешел в нижние мозжечковые ножки. Дорсальнее пирамид между швом и нижним оливным ядром отчетливо видна медиальная петля, кзади от которой находятся медиальный продольный пучок и задний продольный пучок. Снаружи к нижней ножке мозжечка и волокнам слухового нерва прилежит заднее улитковое ядро. Кпереди от входа этого нерва в мозг располагается переднее улитковое ядро. Мост мозга Мост мозга (pons cerebri) является продолжением продолговатого мозга в оральном направлении. Наиболее характерной структурой моста являются его ядра, волокна трапециевидного тела и мостомозжечковые волокна, которые занимают его основание. Скопления волокон моста, поднимающихся в мозжечок, образуют среднюю мозжечковую ножку. По гистологическому строению мост делится на переднюю (базальную) и заднюю часть, или покрышку (tegmentum). Передняя часть моста состоит из идущих в продольном направлении пучков пирамидных волокон и волокон других нисходящих путей, разделенных ядрами моста и поперечными волокнами. В отличие от продолговатого мозга волокна пирамидного пути в мосту представлены некомпактными образованиями, так как идущие поперечно волокна моста разделяют пирамидный путь на отдельные пучки волокон. Из основания моста между продолговатым мозгом и мостом выходит отводящий нерв (nucl. n. abducens, VI), который находится вблизи от средней линии. Латеральнее выходит лицевой нерв (n. facialis, VII), еще латеральнее - преддверно-улитковый нерв (n. vestibulocochlearis, VIII). В боковых отделах моста, в средних его частях, выходит тройничный нерв (n. trigeminus, V). Покрышка моста, так же как и продолговатый мозг, содержит ядра черепных нервов, релейные ядра, ядра ретикулярной формации, а также ядра, осуществляющие взаимосвязь образований различного уровня мозгового ствола. В области перехода от продолговатого мозга к мосту и в нижнем отделе моста располагаются вестибулярные (nucl. vestibularis, VIII) и улитковые ядра (nucl. cochlearis, VIII), которые занимают самые латеральные отделы на поперечном срезе. В вентролатеральных отделах покрышки каудально находится крупное ядро лицевого нерва. В среднем отделе моста, непосредственно у дна IV желудочка, расположено ядро отводящего нерва (nucl. n. abducens, VI) (рис. 2.12). В среднем и верхнем отделах моста вентролатерально, несколько оральнее окончания ядра лицевого нерва, лежат двигательное и чувствительное ядра тройничного нерва.
1 - ядра и волокна моста; 2 - медиальная петля; 3 - медиальное вестибулярное ядро; 4 - дорсальное кохлеарное ядро; 5 - средняя мозжечковая ножка; 6 - ядро лицевого нерва; 7 - верхнее оливное ядро Средний отдел моста занят ядрами ретикулярной формации, которые в продольном направлении могут быть разделены на оральное и каудальные, а каждое из них в поперечном направлении - на медиальную и латеральную части. Медиальные отделы ретикулярных ядер моста содержат более крупные клетки. Медиальная петля меняет свое расположение: она сдвигается вентральнее, а затем вентролатеральнее. В среднем и верхнем отделах моста к медиальной петле примыкают волокна латеральной петли. На границе медиальной и латеральной петель идет латеральный спинно-таламический путь. Кзади от латеральной петли лежит передний спинно-мозжечковый путь, кнутри от него - красноядерно-спинномозговой путь. В дорсальных отделах покрышки у средней линии располагаются волокнистые системы, осуществляющие взаимосвязь образований мозгового ствола. К этим системам относятся медиальный продольный пучок, задний продольный пучок, круглый пучок и др. Среди образований моста следует отметить верхнее оливное ядро, а также переднее и заднее ядра трапециевидного тела, имеющего отношение к восприятию слуха. Оральнее, в верхних и дорсолатеральных отделах моста, находятся верхние мозжечковые ножки, которые образуют боковую стенку IV желудочка. На этих же уровнях расположены верхние вестибулярные ядра и появляются клетки двигательного ядра тройничного нерва. Через мост проходит ряд проводящих путей, которые связывают вышележащие отделы головного мозга со спинным мозгом. Наиболее крупным из них является пирамидный путь, который идет в вентральных отделах моста. Через мост проходят также красноядерно-спинномозговой путь, берущий начало в среднем мозге, и покрышечно-спинномозговой путь, начинающийся в покрышке среднего мозга. В мосту мозга берут начало нисходящие преддверно-спинномозговой путь, идущий из вестибулярных ядер, ретикулярно-спинномозговой путь, берущий начало главным образом в гигантоклеточном ядре и парагигантоклеточных дорсальном и вентральном ядрах ретикулярной формации, а также ряд других систем волокон. Волокна большинства как нисходящих, так и восходящих проводящих путей, идущих через мост мозга, отдают на этом уровне коллатерали, оканчивающиеся на клетках ретикулярной формации. Средний мозг Средний мозг (mesensephalon) является продолжением моста. На базальной поверхности головного мозга средний мозг четко отделяется от моста благодаря поперечным волокнам. С дорсальной поверхности средний мозг отграничивается от моста мозга по уровню перехода IV желудочка в водопровод и расположению нижних холмиков покрышки. На уровне перехода IV желудочка в водопровод верхнюю часть IV желудочка формирует верхний мозговой парус, где образуют перекрест волокна блокового нерва и переднего спинно-мозжечкового пути. В латеральных отделах в средний мозг входят верхние мозжечковые ножки, которые, постепенно погружаясь в него, образуют перекрест у средней линии. Дорсальная часть среднего мозга, расположенная кзади от водопровода, представлена крышей (tectum mesencephali) с ядрами нижних и верхних холмиков. Строение ядер нижних холмиков простое: они состоят из относительно гомогенной массы нервных клеток среднего размера, участвуют в реализации функции слуха и сложных рефлексов на звуковые раздражения. Ядра верхних холмиков организованы более сложно и имеют слоистое строение, принимая участие в осуществлении безусловных рефлексов на зрительные раздражения. Кроме того, эти ядра координируют движения туловища, мимические реакции, движения глазных яблок, головы и т.д. Данные рефлекторные реакции осуществляются благодаря покрышечно-спинномозговому и покрышечно-бульбарному путям. Вентральнее верхних и нижних холмиков крыши находится водопровод среднего мозга, окруженный центральным серым веществом (околоводопроводное серое вещество). В нижнем отделе покрышки среднего мозга на уровне нижних холмиков четверохолмия располагается ядро блокового нерва (nucl. n. trochlearis, IV), состоящее из немногочисленных крупных многоугольных клеток. На уровне среднего и верхнего отделов покрышки локализуется комплекс ядер глазодвигательного нерва, в который входят главное ядро глазодвигательного нерва, крупноклеточное, сходное по морфологии с ядрами блокового и отводящего нервов, мелкоклеточное непарное центральное ядро и наружное мелкоклеточное добавочное ядро. Ядра глазодвигательного нерва располагаются в покрышке среднего мозга у средней линии, вентрально от водопровода, на уровне верхних холмиков крыши среднего мозга. Важными образованиями среднего мозга являются парные красные ядра и черное вещество (рис. 2.13). Красное ядро (nucl. ruber) располагается вентролатеральнее центрального серого вещества среднего мозга. В красном ядре оканчиваются волокна от зубчатых ядер мозжечка и стриопаллидарной системы, начинаются волокна красноядерноспинномозгового, красноядерно-оливного путей, а также волокна, идущие в кору большого мозга. Вентральнее от красного ядра располагается черное вещество (subst. nigra), которое как бы отделяет покрышку среднего мозга от его основания. Основание ножки мозга состоит из волокон, которые связывают кору большого мозга и другие образования конечного мозга с нижележа-
1 - нижний холмик пластинки крыши; 2 - красное ядро; 3 - черное вещество; 4 - ядро глазодвигательного нерва; 5 - основание ножки мозга щими образованиями ствола и спинного мозга. Большая часть основания занята волокнами пирамидного пути. В медиальной части ножки мозга располагаются волокна, идущие из лобных долей к ядрам моста и продолговатого мозга, в средней части - волокна пирамидного пути, в латеральной части - волокна, идущие из височной, затылочной и теменной долей к ядрам моста. Ретикулярная формация Ретикулярная формация (formatio reticularis) получила название в связи с тем, что в ней нервные клетки располагаются на значительном расстоянии друг от друга и разделены большим количеством волокон, идущих во всех направлениях (рис. 2.14). Дендриты нейронов ретикулярной формации тянутся на большом протяжении и дают сравнительно небольшое количество ветвей. Ретикулярная формация проходит через продолговатый мозг, мост, средний мозг и заканчивается в неспецифических ядрах таламуса. На уровне продолговатого мозга и моста ретикулярную формацию можно разделить на латеральную и медиальную части. Медиальная часть содержит более крупные клетки, что позволяет выделить в ретикулярной формации крупноклеточное вентромедиальное и мелкоклеточное дорсолатеральное ядра ствола мозга. В области перехода продолговатого мозга в мост в вентромедиальных отделах ретикулярной формации располагаются гигантоклеточное ретикулярное ядро, парагигантоклеточное дорсальное и латеральное ретикулярные ядра. Дорсолатеральную часть ретикулярной формации на этом уровне занимает мелкоклеточное ретикулярное ядро. Ретикулярная формация играет важную роль в осуществлении взаимосвязи между проводящими путями и ядерными образованиями ствола мозга, а также в регуляции функций надстволовых отделов
1 - восходящая активирующая ретикулярная формация ствола мозга; 2 - гипоталамус; 3 - таламус; 4 - кора большого мозга; 5 - мозжечок; 6 - афферентные коллатерали; 7 - продолговатый мозг; 8 - мост мозга; 9 - средний мозг ЦНС. К ней дают коллатерали все основные восходящие и нисходящие проводники. Ретикулярная формация имеет взаимные связи с гипоталамусом, таламусом, стриопаллидарной системой, корой больших полушарий (особенно в области лимбической системы), а также с мозжечком, ядрами ствола мозга, системой заднего (медиального) продольного пучка. Волокна, соединяющие ядра ретикулярной формации с системой заднего продольного пучка, проходящего вблизи средней линии у дна IV желудочка, связывают в единую сеть ядра верхних и нижних бугорков четверохолмия, всех глазодвигательных черепных нервов, вестибулярных, добавочных, блуждающих нервов, создавая основу для реализации безусловных рефлексов. Пути, идущие от ретикулярной формации к спинному мозгу (ретикулоспинальные), участвуют в регуляции мышечного тонуса. Ядра ретикулярной формации на уровне моста и перехода от моста к продолговатому мозгу участвуют в регуляции дыхания и сердечно-сосудистой деятельности, реализуя свои влияния через связи с ядрами блуждающего нерва и некоторых других систем мозга. Ретикулярная формация оказывает активирующее действие на все вышележащие отделы головного мозга и обеспечивает, в частности регуляцию сна и бодрствования. Мозжечок Мозжечок (cerebellum) расположен в задней черепной ямке, кзади от продолговатого мозга и моста, под затылочными долями полушарий большого мозга (рис. 2.15). От него мозжечок отделяется твердой оболочкой головного мозга - наметом мозжечка (tentorium cerebelli). Мозжечок состоит из средней части, или червя (vermis cerebelli), и двух боковых частей - полу-шарий (hemispherium cerebelli). В мозжечке различают переднюю, верхнюю и нижнюю поверхности и передний и задний края. Принято деление мозжечка на переднюю, заднюю, и клочково-узелковую доли, а термины «червь» и «полушария» используются при определении центральных или периферических отделов мозжечка. Кора мозжечка имеет трехслойное строение: наружный (молекулярный) содержит небольшое количество малых звездчатых клеток; внутренний (зернистый) образован тесно прилежащими друг к другу клетками-зернами и более редкими крупными звездчатыми нейронами, между ними располагается слой крупных клеток Пуркинье (рис. 2.16). В белом веществе мозжечка расположены 4 пары ядер. Наиболее поверхностно (в полушарии мозжечка) находится крупное зубчатое ядро (nucl. dentatus). Кнутри от него расположено пробковидное ядро (nucl. embo-
1 - бугор червя; 2 - горизонтальная щель; 3 - пирамида червя; 4 - миндалина мозжечка; 5 - клочок; 6 - ножка клочка; 7 - преддверно-улитковый нерв; 8 - промежуточный нерв; 9 - лицевой нерв; 10 - тройничный нерв; 11 - корешки языкоглоточного и блуждающего нервов; 12 - отводящий нерв; 13 - корешки подъязычного нерва; 14 - базилярная борозда; 15 - мост мозга; 16 - пирамида продолговатого мозга; 17 - олива liformis). Ядро шатра (nucl. fastigii) лежит в белом веществе червя, у средней линии, над IV желудочком, а между ним и пробковидным ядром находится шаровидное ядро (nucl. globosus) (рис. 2.17). В мозжечке различают экстра- и эндоцеребеллярную системы волокон. К экстрацеребеллярной системе относятся длинные проекционные афферентные и эфферентные волокна, связывающие мозжечок с другими отделами головного и спинного мозга. Эндоцеребеллярная система состоит из ассоциативных, комиссуральных и коротких проекционных волокон. Ассоциативные волокна связывают отдельные участки коры мозжечка между собой, комиссуральные - противоположные полушария мозжечка, а короткие проекционные волокна - кору мозжечка с его ядрами. Связи мозжечка с другими отделами ЦНС осуществляются посредством трех пар мозжечковых ножек: нижней, средней и верхней. Нижняя и средняя мозжечковые ножки состоят преимущественно из афферентных путей, а верхняя мозжечковая ножка принадлежит в основном эфферентной системе. Мозжечок осуществляет постоянный контроль двигательной активности. Он участвует в координации движений, регуляции мышечного тонуса, сохранении позы и равновесия тела. Промежуточный мозг Промежуточный мозг (diencephalon) находится между средним мозгом и полушариями большого мозга, включает III желудочек и образования, формирующие его стенки. В промежуточном мозге выделяют верхний отдел - эпиталамус, средний отдел - таламус, нижний отдел - гипоталамус и задний отдел - метаталамус. III желудочек имеет форму
1 - вьющиеся волокна; 2 - аксон клетки Пуркинье; 3 - клубочек; 4 - клетка Пуркинье; 5 - наружная звездчатая клетка; 6 - параллельные волокна; 7 - молекулярный слой; 8 - слой клетки Пуркинье; 9 - гранулярный слой; 10 - белое вещество; 11 - мшистые волокна
узкой щели, его дно образовано гипоталамусом. Переднюю стенку III желудочка составляет тонкая концевая пластинка, которая начинается у зрительного перекреста и переходит в ростральную пластинку мозолистого тела. В верхнем отделе передней стенки III желудочка находятся столбы свода. Около столбов свода в его передней стенке находится отверстие, соединяющее III желудочек с парными боковыми желудочками (отверстие Монро). Боковые стенки III желудочка образованы таламусом. Под задней спайкой мозга III желудочек переходит в водопровод среднего мозга. Таламус (thalamus) имеет сложное цитоархитектоническое стро- ение. Внутренняя поверхность таламуса отделяется от верхней мозговой полоской. Верхнюю поверхность покрывает белое вещество. Передняя часть верхней поверхности утолщается и образует передний бугорок (tuberculum thalami anterius), а задний бугорок образует подушку (pulvinar). Латерально верхняя поверхность таламуса граничит с парным хвостатым ядром (nucl. caudatus), отделяясь от него пограничной полоской. Наружная поверхность таламуса отделяется внутренней капсулой от чечевицеобразного ядра и головки хвостатого ядра. Среди множества ядер таламуса выделяют следующие группы: • комплекс специфических, или релейных, таламических ядер, через которые проводится афферентная информация определенной модальности; • неспецифические таламические ядра, не связанные с проведением афферентной информации и проецирующиеся на кору больших полушарий более диффузно, чем специфические ядра; • ассоциативные ядра таламуса, получающие раздражения от других ядер таламуса и передающие эту информацию на ассоциативные области коры головного мозга. Основные ядра таламуса: • передние (nucll. anterior); • срединные (nucll. mediales); • дорсолатеральные (nucll. dorsolateralis); • внутрипластинчатые (nucll. intralaminares); • вентролатеральные (nucll. ventrolaterales); • задние (nucll. posteriores); • ретикулярные (nucll. reticulares). Подбугорное ядро (nucl. subthalamicus) относится к субталамической области промежуточного мозга и состоит из однотипных мультиполярных клеток. К субталамической области относятся также ядра Н-, Н1- и Н2-полей и неопределенная зона (zona incerta). Поле Н1 располагается под таламусом и состоит из волокон, соединяющих гипоталамус с полосатым телом. Под полем Н1 находится неопределенная зона, переходящая в перивентрикулярную зону III желудочка. Под неопределенной зоной лежит поле Н2, соединяющее бледный шар с подбугорным ядром и перивентрикулярными ядрами гипоталамуса. К эпиталамусу относятся поводки, спайка поводков, задняя спайка и шишковидное тело. В треугольнике поводка располагаются ядра поводка: медиальное, состоящее из мелких клеток, и латеральное, в котором преобладают крупные клетки. К метаталамусу относятся медиальное и латеральное коленчатые тела. Латеральное коленчатое тело находится под подушкой таламуса, оно является одним из основных подкорковых центров для передачи зрительных ощущений, а также участвует в осуществлении бинокулярного зрения. Медиальное коленчатое тело располагается между верхними бугорками четверохолмия и подушкой таламуса. В медиальном коленчатом теле выделяют дорсальное и вентральное ядра. На клетках медиального коленчатого тела оканчиваются волокна латеральной петли и берет начало центральный слуховой путь, идущий к слуховой коре. Медиальное коленчатое тело является подкорковым центром слухового анализатора. Гипоталамус (hypothalamus) - филогенетически наиболее старая часть промежуточного мозга. Гипоталамус имеет сложное строение (рис. 2.18). В преоптической (передней гипоталамической) области выделяются медиальное и латеральное преоптические ядра, паравентрикулярное и супраоптическое ядра, переднее гипоталамическое и супрахиазмальное ядра. В медиальной части промежуточной гипоталамической области находятся дорсомедиальное и вентромедиальное гипоталамические ядра, ядро воронки, которое также называют дугообразным ядром. В латеральной части промежуточной области гипоталамуса расположены латеральное гипоталамическое, серобугорное, серобугорно-сосцевидное и перифорникальное ядра. Задняя гипоталамическая область содержит медиальное и латеральное ядра сосцевидного тела, заднее гипоталамическое ядро. Гипоталамус обладает сложной системой афферентных и эфферентных путей. Афферентные пути: 1) медиальный пучок переднего мозга, связывающий перегородку и преоптическую область с ядрами гипоталамуса; 2) свод, соединяющий кору гиппокампа с гипоталамусом; 3) таламогипофизарные волокна, соединяющие таламус с гипоталамусом; 4) покрышечно-сосцевидный пучок, содержащий волокна, идущие из среднего мозга к гипоталамусу; 5) задний продольный пучок, несущий импульсы от ствола мозга к гипоталамусу; 6) паллидогипоталамический путь. Установлены также непрямые мозжечково-гипоталамические связи, оптико-гипоталамические пути, ваго-супраоптические связи.
А - Различные срезы: 1 - передний (ростральный) отдел гипоталамуса; 2 - передняя комиссура; 3 - паравентрикулярное ядро; 4 - супрахиазматическое ядро; 5 - супраоптическое ядро; 6 - зрительный перекрест; 7 - свод; 8 - зрительный тракт; 9 - латеральное гипоталамическое поле; 10 - дорсомедиальное гипоталамическое ядро; 11 - вентромедиальное гипоталамическое ядро; 12 - туберомамиллярное ядро; 13 - инфундибулярное ядро; 14 - туберальное латеральное ядро; 15 - латеральное мамиллярное ядро; 16 - медиальное мамиллярное ядро; 17 - ножки мозга. Б - Сагиттальный срез: 1 - паравентрикулярное ядро; 2 - сосцевидно-таламический пучок; 3 - дорсомедиальное гипоталамическое ядро; 4 - вентромедиальное гипоталамическое ядро; 5 - мост мозга; 6 - супраоптический гипофизарный путь; 7 - нейрогипофиз; 8 - гипофиз; 9 - аденогипофиз; 10 - зрительный перекрест; 11 - супраоптическое ядро; 12 - преоптическое ядро Эфферентные пути гипоталамуса: 1) пучки волокон перивентрикулярной системы к заднемедиальным таламическим ядрам и преимущественно к нижней части ствола мозга, ретикулярной формации среднего мозга и спинному мозгу; 2) сосцевидные пучки, идущие к передним ядрам таламуса и ядрам среднего мозга; 3) гипоталамо-гипофизарный путь к нейрогипофизу. Кроме того, имеется комиссуральный путь, благодаря которому медиальные гипоталамические ядра одной стороны вступают в контакт с медиальными и латеральными ядрами другой. Гипоталамус является одним из основных образований мозга, участвующих в регуляции вегетативных, трофических и нейроэндокринных функций, он играет существенную роль в регуляции и интеграции деятельности внутренних органов, желез внутренней секреции, симпатического и парасимпатического отделов вегетативной нервной системы. Гипоталамусу свойственна очень важная нейросекреторная функция, в частности регуляции выделения гормонов гипофизом. В нервных клетках гипоталамических ядер образуется нейросекрет, причем нейросекреторные гранулы, вырабатывающиеся в разных ядрах, различаются по химическому составу и свойствам. Регулирующие влияния гипоталамуса передаются к эндокринным железам не только с помощью гипоталамических нейрогормонов (рилизинг-факторов), переносимых с током крови и действующих гуморально, но и по эфферентным нервным волокнам. Одной из функций гипоталамической области является регуляция деятельности сердечно-сосудистой системы. При нарушении функций гипоталамических ядер происходит изменение терморегуляции и трофики тканей. Гипоталамус участвует в формировании биологических мотиваций и эмоций. Базальные ядра Базальные ядра (nucll. basales) - скопления серого вещества в глубине полушарий большого мозга (рис. 2.19, 2.20). К ним относятся хвостатое ядро, скорлупа, бледный шар, ограда и миндалевидное тело. Хвостатое ядро (nucl. caudatus) представляет собой крупное образование грушевидной формы, расположенное кпереди и кнутри от таламуса и отделенное от него внутренней капсулой (capsula interna). Ростральная, утолщенная часть носит название головки хво-статого ядра (caput nucl. caudati), кзади от нее хвостатое ядро суживается и образует тело хвоста- того ядра (corpus nucl. caudati). Истонченный задний отдел, обозначаемый как хвост (cauda nucl. caudati), загибается в височную долю, где сливается с миндалевидным телом. Верхняя и внутренняя поверхности хвостатого ядра образуют стенку бокового желудочка. От скорлупы (putamen) головка хвостатого ядра отделяется передней ножкой внутренней капсулы. Эти два образования соединены друг с другом клеточными мостиками и местами идентичны по структуре. Основную популяцию составляют пирамидные, звездчатые и веретенообразные нейроны, преобладают мелкие или средние клетки при небольшом содержании нейронов крупного размера. Скорлупа вместе с наружным и внутренним сегментами бледного шара объединяется под общим названием «чечевицеобразное ядро» (nucl. lentiformis), но отличается более плотным расположением клеток. Сходство в клеточном строении, развитии и функциях хвостатого ядра и скорлупы позволило их объединить под названием «полосатое тело» (corpus striatum). Бледный шар (globus pallidus) в отличие от хвостатого ядра и скорлупы макроскопически имеет очень бледный цвет из-за большого количества миелиновых волокон. Бледный шар отличается от структур полосатого тела особенностями эволюционного развития (он появляется в фило-
1 - продольная щель большого мозга; 2 - свод; 3 - мозолистое тело; 4 - сосудистое сплетение бокового желудочка; 5 - лучистость мозолистого тела; 6 - прецентральная извилина; 7 - центральная борозда; 8 - постцентральная извилина; 9 - медиальное ядро таламуса; 10 - хвост хвостатого тела; 11 - задняя часть передней спайки; 12 - гиппокамп; 13 - неопределенная зона; 14 - субталамическое ядро; 15 - III желудочек; 16 - сосцевидные тела; 17 - сосцевидно-таламический пучок; 18 - основание ножки мозга; 19 - миндалевидное тело; 20 - зрительный тракт; 21 - нижний рог бокового желудочка; 22 - верхняя височная борозда; 23 - ограда; 24 - островок; 25 - латеральная борозда; 26 - покрышка; 27 - скорлупа; 28 - бледный шар; 29 - внутренняя капсула; 30 - латеральные ядра таламуса; 31 - хвостатое ядро; 32 - мозговая пластинка таламуса; 33 - передние ядра таламуса
Наибольшее число афферентных связей хвостатое ядро, скорлупа и бледный шар получают из коры, особенно из ее передних отделов, моторной и соматосенсорной зон. Вторым важнейшим источником афферентации этих образований являются пути, идущие от таламуса, в основном от неспецифических внутрипластинчатых и срединных ядер. Источником афферентных связей бледного шара является также средний мозг, прежде всего черное вещество. Особенно много волокон из черного вещества поступает в хвостатое ядро, а нигростриарный дофаминергический путь имеет большое функциональное значение. Имеются афферентные связи базальных ганглиев с миндалевидными телами, ретикулярной формацией, гиппокампом, мозжечком. Эфферентные связи осуществляются через пути из бледного шара в таламус. Проекции хвостатого ядра и скорлупы на таламус в основном не прямые, а опосредованные через бледный шар. Известны также восходящие проекции от паллидума к коре, главным образом к лобным долям ипсилатерального полушария. При этом установлено перекрытие проекций волокон из хвостатого ядра, скорлупы и бледного шара в ряде полей коры большого мозга. Существуют и каудато-паллидарные связи, а также проекции стриатума и паллидума на черное вещество, ретикулярную формацию и дру- Рис. 2.20. Горизонтальный срез большого мозга на уровне мозолистого тела. 1 - колено мозолистого тела; 2 - свод; 3 - наружная капсула; 4 - самая наружная капсула; 5 - ограда; 6 - чечевицеобразное ядро; 7 - III желудочек; 8 - внутренняя капсула; 9 - сосудистое сплетение бокового желудочка; 10 - задняя таламическая лучистость; 11 - шпорная борозда; 12 - продольная щель большого мозга; 13 -валик мозолистого тела; 14 - задний рог бокового желудочка; 15 - латеральные ядра таламуса; 16 - медиальные ядра таламуса; 17 - передние ядра таламуса; 18 - островок; 19 - внутренняя капсула (передняя ножка); 20 - головка хвостатого ядра; 21 - передний рог бокового желудочка гие системы мозга. Сложностью организации и богатством афферентных и эфферентных связей определяется полифункциональность структур мозга, входящих в стриопаллидарную систему. Все эти структуры играют важную роль в контроле двигательных реакций, реализации условно-рефлекторной деятельности и возникновении сложных форм поведенческих реакций. Ограда (claustrum) представляет собой узкую пластинку серого вещества, которая располагается латеральнее чечевицеобразного ядра и отделена от него наружной капсулой. Миндалевидное тело (corpus amigdaloideum) располагается в передневерхней части парагиппокампальной извилины и в этой зоне соприкасается с древней корой. Миндалевидное тело содержит две группы ядер: одна из них состоит из базального и латерального ядер, другая - из кортикального, медиального, центрального ядер и ядра латерального обонятельного тракта. Эти ядра различаются по гистологическим и цитоархитектоническим особенностям. Миндалевидное тело принимает участие в корригирующем влиянии на деятельность стволовых образований, оно связано с широким диапазоном поведенческих, эмоциональных, половых, эндокринных, обменных реакций. Внутренняя капсула - плотный слой проекционных (афферентных и эфферентных) волокон, связывающих кору большого мозга с нижележащими отделами ЦНС, ограниченный хвостатым ядром и таламусом с одной стороны, и лентикулярным ядром - с другой. На горизонтальном срезе внутренняя капсула выглядит как тупой угол, открытый кнаружи (см. рис. 2.21). Во внутренней капсуле выделяют переднюю и заднюю ножки и соединяющее их колено. В передней ножке проходят эфферентные волокна, исходящие из коры лобной доли и направляющиеся к таламусу (корково-таламический путь) и через мост к мозжечку (корково-мостомозжечковый путь). Через колено и передние 2/3 задней ножки идут нисходящие пирамидные волокна от двигательной коры (в колене - к двигательным ядрам черепных нервов, корково-ядерная часть пирамидного тракта; в передних 2/3 заднего бедра - к передним рогам спинного мозга, корково-спинномозговая часть пирамидного тракта). В задней трети задней ножки проходят восходящие чувствительные волокна от таламуса к задней центральной извилине (таламокорковый путь), восходящие пути зрительного и слухового анализаторов, направляющиеся соответственно к затылочной и височной долям, а также нисходящие эфферентные волокна, идущие от нижних отделов височной и затылочной долей через мост к мозжечку (затылочно-височно-мостомозжечковый путь).
1 - корково-таламический путь, 2 - корково-мосто-мозжечковый путь, 3 - корково-ядерная часть пирамидного тракта, 4 - корково-спинномозговая часть пирамидного тракта, 5 - таламокорковый путь, 6 - путь зрительного анализатора, 7 - путь слухового анализатора, 8 - затылочно-височно-мостомозжечковый путь Кора большого мозга Полушария большого мозга (рис. 2.22) разделены глубокой бороздой, которая доходит до мозолистого тела - массивного слоя волокон, соединяющих оба полушария. Каждое полушарие имеет три полюса: лобный, затылочный и височный. Макроскопически в каждом полушарии выделяются лобная, теменная, затылочная, височная доли и островок. Поверхность большого мозга образована корой, состоящей из нервных клеток. Под корой большого мозга располагается слой волокон, связывающих кору большого мозга с подкорковыми образованиями и отдельные ее поля между собой. Поверхность полушарий большого мозга изрезана бороздами, разделяющими ее на доли и извилины. Различают первичные борозды, встречающиеся постоянно, глубокие, рано появляющиеся на мозге эмбриона; вторичные, тоже постоянные, но появляющиеся позднее, и третичные, непостоянные борозды. Среди наиболее крупных пер- вичных борозд выделяют: 1) центральную (роландову) борозду (sulcus centralis), которая отделяет лобную долю от теменной; 2) латеральную (сильвиеву) борозду (sulcus lateralis), которая отделяет лобную и теменную доли от височной; 3) теменно-затылочную борозду (sulcus parietooccipitalis), отделяющую теменную долю от затылочной. Первые две борозды располагаются на наружной поверхности полушарий, третья - на медиальной. Дата добавления: 2015-12-16 | Просмотры: 4889 | Нарушение авторских прав |
 Рис. 2.1. Строение спинного мозга. А - Расположение спинного мозга в спинномозговом канале. C - Шейные спинномозговые нервы (СI-СVII, синие); Th - Грудные спинномозговые нервы (ThI-ThXII, фиолетовые); L - Поясничные спинномозговые нервы (LI-LV, бирюзовые); S - Крестцовые спинномозговые нервы (SI-SV, светло-зеленые); Co - Копчиковые спинномозговые нервы (Со, серые); 1 - тело позвонка; 2 - межпоперечная связка; 3 - эпидуральные вены; 4 - реберно-позвоночный сустав; 5 - паутинная оболочка; 6 - ребро; 7 - зубчатая связка; 8 - вентральный корешок; 9 - спинномозговой нерв; 10 - задняя ветвь (кожа и мышцы спины); 11 - эпидуральное пространство; 12 - мягкая мозговая оболочка; 13 - субарахноидальное пространство; 14 - спинальный ганглий; 15 - корешковый рукав; 16 - межпозвонковое отверстие; 17 - менингеальная ветвь; 18 - задняя продольная связка; 19 - твердая мозговая оболочка; 20 - межпозвоночный диск, фиброзное кольцо; 21 - атлант; 22 - крестец; 23 - копчик. Б - Сегменты спинного мозга и спинномозговые нервы
Рис. 2.1. Строение спинного мозга. А - Расположение спинного мозга в спинномозговом канале. C - Шейные спинномозговые нервы (СI-СVII, синие); Th - Грудные спинномозговые нервы (ThI-ThXII, фиолетовые); L - Поясничные спинномозговые нервы (LI-LV, бирюзовые); S - Крестцовые спинномозговые нервы (SI-SV, светло-зеленые); Co - Копчиковые спинномозговые нервы (Со, серые); 1 - тело позвонка; 2 - межпоперечная связка; 3 - эпидуральные вены; 4 - реберно-позвоночный сустав; 5 - паутинная оболочка; 6 - ребро; 7 - зубчатая связка; 8 - вентральный корешок; 9 - спинномозговой нерв; 10 - задняя ветвь (кожа и мышцы спины); 11 - эпидуральное пространство; 12 - мягкая мозговая оболочка; 13 - субарахноидальное пространство; 14 - спинальный ганглий; 15 - корешковый рукав; 16 - межпозвонковое отверстие; 17 - менингеальная ветвь; 18 - задняя продольная связка; 19 - твердая мозговая оболочка; 20 - межпозвоночный диск, фиброзное кольцо; 21 - атлант; 22 - крестец; 23 - копчик. Б - Сегменты спинного мозга и спинномозговые нервы Рис. 2.2. Поперечный срез спинного мозга (схема).
Рис. 2.2. Поперечный срез спинного мозга (схема). Рис. 2.3. Поперечный срез спинного мозга на уровне верхнегрудного отдела. Проводящие пути (схема). 1 - задняя срединная перегородка; 2 - тонкий пучок; 3 - клиновидный пучок; 4 - задний рог; 5 - задний спинно-мозжечковый путь; 6 - центральный канал; 7 - боковой рог; 8 - латеральный спинно-таламический путь; 9 - передний спинно-мозжечковый путь; 10 - передний спинно-таламический путь; 11 - передний рог; 12 - передняя срединная щель; 13 - оливоспинномозговой путь; 14 - передний корково-спинномозговой (пирамидный) путь; 15 - передний ретикулярно-спинномозговой путь; 16 - преддверно-спинномозговой путь; 17 - бульборетикулярно-спинномозговой путь; 18 - передняя белая спайка; 19 - серая спайка; 20 - красноядерно-спинномозговой путь; 21 - латеральный корково-спинномозговой (пирамидный) путь; 22 - задняя белая спайка; 23 - грудной столб (столб Кларка)
Рис. 2.3. Поперечный срез спинного мозга на уровне верхнегрудного отдела. Проводящие пути (схема). 1 - задняя срединная перегородка; 2 - тонкий пучок; 3 - клиновидный пучок; 4 - задний рог; 5 - задний спинно-мозжечковый путь; 6 - центральный канал; 7 - боковой рог; 8 - латеральный спинно-таламический путь; 9 - передний спинно-мозжечковый путь; 10 - передний спинно-таламический путь; 11 - передний рог; 12 - передняя срединная щель; 13 - оливоспинномозговой путь; 14 - передний корково-спинномозговой (пирамидный) путь; 15 - передний ретикулярно-спинномозговой путь; 16 - преддверно-спинномозговой путь; 17 - бульборетикулярно-спинномозговой путь; 18 - передняя белая спайка; 19 - серая спайка; 20 - красноядерно-спинномозговой путь; 21 - латеральный корково-спинномозговой (пирамидный) путь; 22 - задняя белая спайка; 23 - грудной столб (столб Кларка)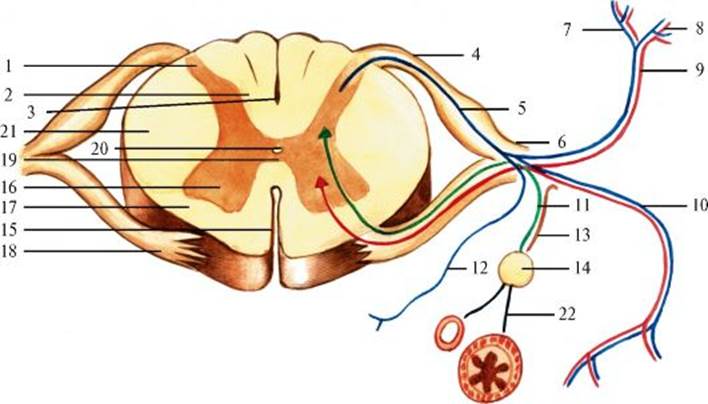 Рис. 2.4. Спинномозговой нерв.
Рис. 2.4. Спинномозговой нерв. Рис. 2.5. Шейное сплетение.
Рис. 2.5. Шейное сплетение. Рис. 2.6. Плечевое сплетение.
Рис. 2.6. Плечевое сплетение. Рис. 2.7. Поясничное сплетение. 1 - подвздошно-подчревный нерв (ThXII; LI); 2 - подвздошная ветвь; 3 - подчревная ветвь; 4 - подвздошнопаховый нерв (LI); 5 - бедренно-половой нерв (LI, LII); 6 - бедренная ветвь; 7 - половая ветвь; 8 - латеральный кожный нерв бедра (LII, LIII); 9 - мышечные ветви бедренного нерва к подвздошным мышцам; 10 - бедренный нерв (LII-LIV); 11 - запирательный нерв (LII-LIV); 12 - ветвь к крестцовому сплетению; 13 - задние двигательные ветви к межпоперечным мышцам поясницы и квадратной мышце поясницы (обозначены звездочками)
Рис. 2.7. Поясничное сплетение. 1 - подвздошно-подчревный нерв (ThXII; LI); 2 - подвздошная ветвь; 3 - подчревная ветвь; 4 - подвздошнопаховый нерв (LI); 5 - бедренно-половой нерв (LI, LII); 6 - бедренная ветвь; 7 - половая ветвь; 8 - латеральный кожный нерв бедра (LII, LIII); 9 - мышечные ветви бедренного нерва к подвздошным мышцам; 10 - бедренный нерв (LII-LIV); 11 - запирательный нерв (LII-LIV); 12 - ветвь к крестцовому сплетению; 13 - задние двигательные ветви к межпоперечным мышцам поясницы и квадратной мышце поясницы (обозначены звездочками) Рис. 2.8. Крестцовое сплетение.
Рис. 2.8. Крестцовое сплетение.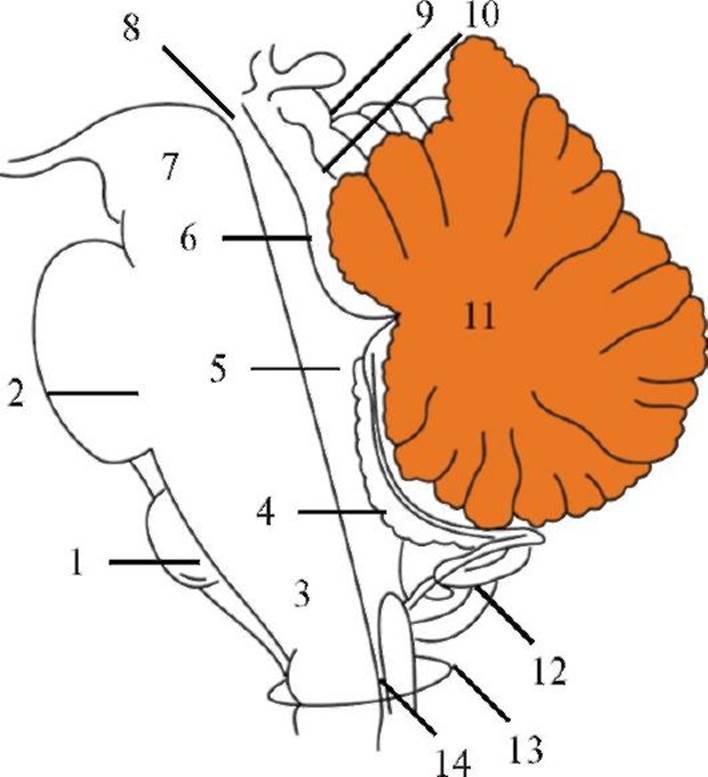 Рис. 2.9. Схема строения ствола мозга (сагиттальный срез). 1 - олива; 2 - мост; 3 - продолговатый мозг; 4 - хориоидальное сплетение; 5 - IV желудочек; 6 - центральный водопровод; 7 - средний мозг; 8 - центральный водопровод; 9 - верхние бугорки; 10 - нижние бугорки; 11 - мозжечок; 12 - миндалина мозжечка; 13 - большое затылочное отверстие; 14 - центральный канал спинного мозга
Рис. 2.9. Схема строения ствола мозга (сагиттальный срез). 1 - олива; 2 - мост; 3 - продолговатый мозг; 4 - хориоидальное сплетение; 5 - IV желудочек; 6 - центральный водопровод; 7 - средний мозг; 8 - центральный водопровод; 9 - верхние бугорки; 10 - нижние бугорки; 11 - мозжечок; 12 - миндалина мозжечка; 13 - большое затылочное отверстие; 14 - центральный канал спинного мозга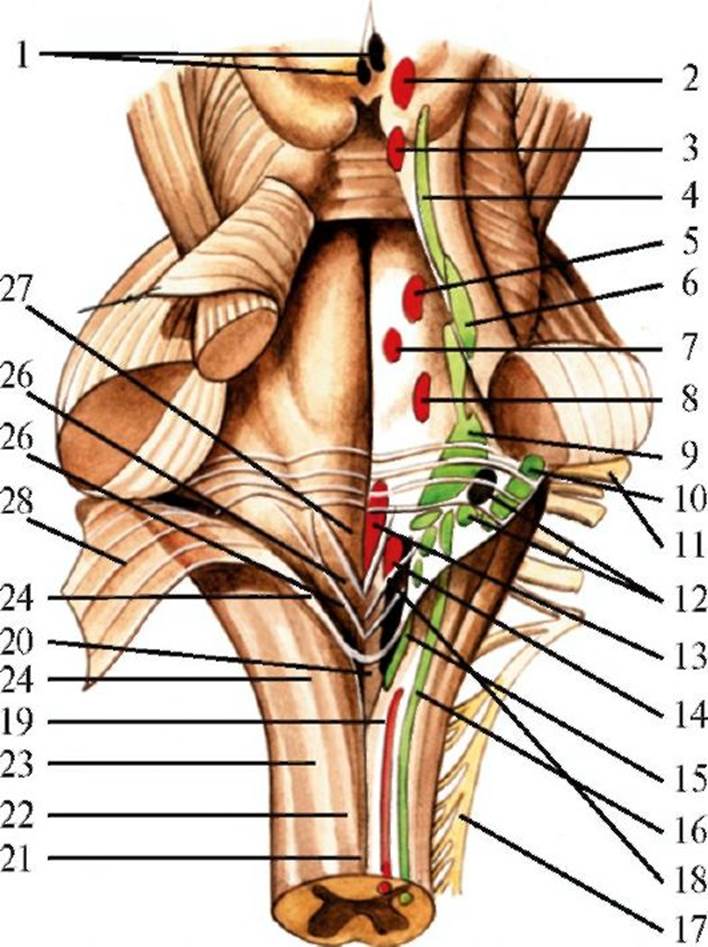 Рис. 2.10. Расположение ядер черепных нервов в стволе мозга. Ромбовидная ямка.
Рис. 2.10. Расположение ядер черепных нервов в стволе мозга. Ромбовидная ямка. Рис. 2.11. Поперечный срез продолговатого мозга (А). Схема (Б). 1 - ядро подъязычного нерва; 2 - заднее ядро блуждающего нерва; 3 - ядро одиночного пути; 4 - добавочное клиновидное ядро; 5 - нижняя мозжечковая ножка; 6 - ядро (нижнее) спинномозгового пути тройничного нерва; 7 - центральное ретикулярное ядро продолговатого мозга (вентральное и дорсальное подъядра); 8 - нижнее оливное ядро; 9 - пирамидный пучок; 10 - медиальная петля
Рис. 2.11. Поперечный срез продолговатого мозга (А). Схема (Б). 1 - ядро подъязычного нерва; 2 - заднее ядро блуждающего нерва; 3 - ядро одиночного пути; 4 - добавочное клиновидное ядро; 5 - нижняя мозжечковая ножка; 6 - ядро (нижнее) спинномозгового пути тройничного нерва; 7 - центральное ретикулярное ядро продолговатого мозга (вентральное и дорсальное подъядра); 8 - нижнее оливное ядро; 9 - пирамидный пучок; 10 - медиальная петля Рис. 2.12. Поперечный срез моста мозга (А). Схема (Б).
Рис. 2.12. Поперечный срез моста мозга (А). Схема (Б).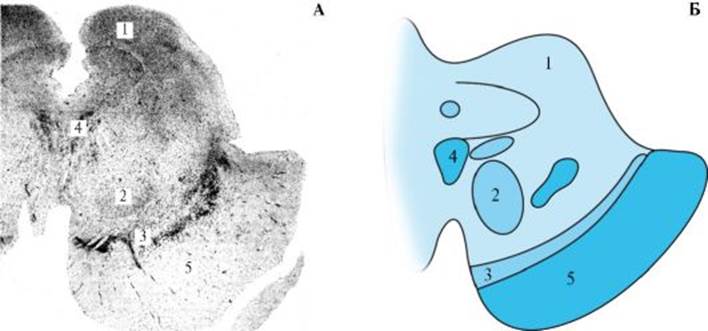 Рис. 2.13. Поперечный срез среднего мозга (А). Схема (Б).
Рис. 2.13. Поперечный срез среднего мозга (А). Схема (Б). Рис. 2.14. Восходящая активирующая ретикулярная формация ствола мозга (схема).
Рис. 2.14. Восходящая активирующая ретикулярная формация ствола мозга (схема). Рис. 2.15. Мозжечок.
Рис. 2.15. Мозжечок. Рис. 2.16. Клеточное строение коры мозжечка.
Рис. 2.16. Клеточное строение коры мозжечка. Рис. 2.17. Ядра мозжечка. 1 - шаровидное ядро; 2 - зубчатое ядро; 3 - пробковидное ядро; 4 - ядро шатра
Рис. 2.17. Ядра мозжечка. 1 - шаровидное ядро; 2 - зубчатое ядро; 3 - пробковидное ядро; 4 - ядро шатра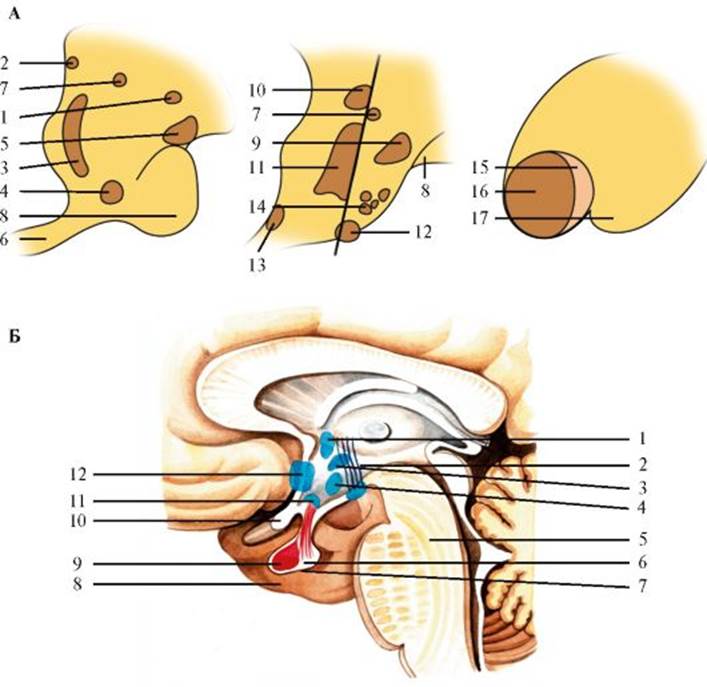 Рис. 2.18. Гипоталамус.
Рис. 2.18. Гипоталамус. Рис. 2.19. Фронтальный срез большого мозга на уровне сосцевидных тел.
Рис. 2.19. Фронтальный срез большого мозга на уровне сосцевидных тел.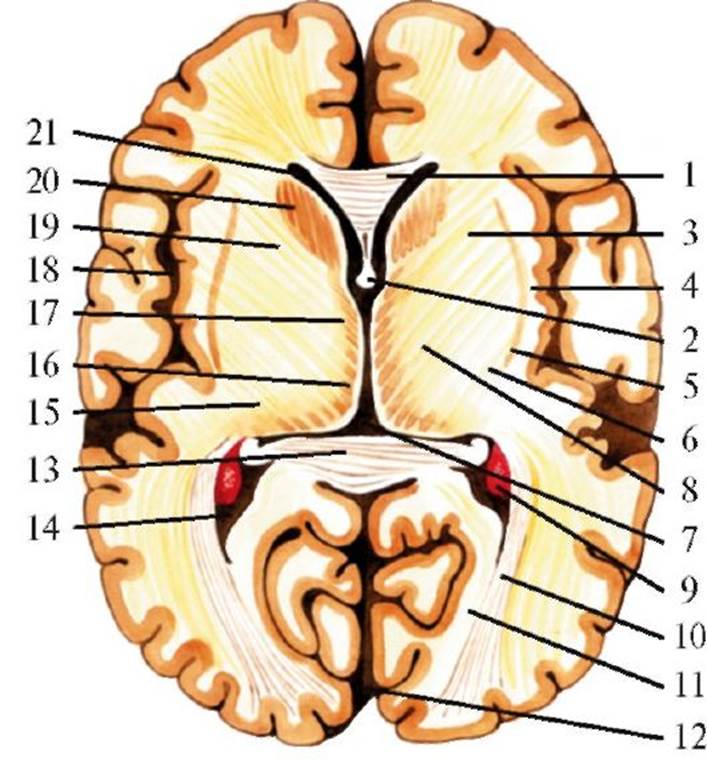 и онтогенезе раньше, чем хвостатое ядро и скорлупа), гистологической структурой и функциями; подразделяется на наружный и внутренний сегменты.
и онтогенезе раньше, чем хвостатое ядро и скорлупа), гистологической структурой и функциями; подразделяется на наружный и внутренний сегменты. Рис. 2.21. Строение внутренней капсулы.
Рис. 2.21. Строение внутренней капсулы.