|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ГЛАВА 3. ЧУВСТВИТЕЛЬНОСТЬ И ЕЕ НАРУШЕНИЯЧувствительность - способность организма воспринимать раздражения, исходящие из окружающей среды или от собственных тканей и органов. Учение И.П. Павлова об анализаторах заложило основы понимания природы и механизмов чувствительности. Каждый анализатор состоит из периферического (рецепторного) отдела, проводниковой части и коркового отдела. Рецепторы - специализированные чувствительные образования, способные воспринимать изменения внутри или вне организма и преобразовывать их в нервные импульсы. Рецепторы представляют собой периферические окончания афферентных нервных волокон, являющихся периферическими отростками псевдоуниполярных нейронов спинномозговых (спинальных) ганглиев. Благодаря специализации рецепторов осуществляется первый этап анализа внешних раздражителей - разложение целого на части, дифференциация характера и качества сигналов. Все виды внешних воздействий (сенсорные стимулы) трансформируются в нервные импульсы, поступающие в ЦНС. Трансформация сенсорного стимула в нервный импульс может происходить путем прямой активации ионных каналов рецепторов (как в случае слухового - звукового раздражения) или путем непрямой активации посредством внутриклеточной системы вторичных мессенджеров (как в случае зрительного раздражения). Во всех случаях нервный импульс проводится в виде потенциала действия, распространяющегося по нервным волокнам от клетки к клетке. Чувствительность имеет специфичность, или модальность, которая определяется прежде всего специализированными структурами, принимающими дифференцированную афферентную информацию (рецепторы). В зависимости от функциональных особенностей рецепторы подразделяются на экстерорецепторы (находятся в кожных покровах и информируют о происходящем в окружающей среде), телерецепторы (органы зрения и слуха), проприорецепторы (обеспечивают информацию о напряжении мышц и сухожилий, движениях и положении частей тела) и интерорецепторы («сообщающие» о состоянии внутри организма). Имеются также осмо-, хемо-, барорецепторы и др. Кожные рецепторы можно классифицировать в соответствии с типом стимула, на который они отвечают, на механорецепторы (реагиру- ющие на прикосновение, давление), терморецепторы (реагирующие на холод, тепло) и ноцицептивные, или болевые, рецепторы (реагирующие на боль). Этих рецепторов много в коже, особенно между эпидермисом и тканью, поэтому кожа может рассматриваться как чувствительный орган, т.е. вся поверхность тела. Механорецепторы реагируют на тактильные стимулы (прикосновение к коже, давление) и бывают быстро или медленно адаптирующимися. Быстро адаптирующимися являются рецепторы волосяных фолликулов, тельца Мейсснера (расположены в коже, лишенной волосяного покрова) и тельца Пачини подкожной ткани (рис. 3.1). Рецепторы волосяных фолликулов и тельца Мейсснера отвечают на стимулы, поступающие с частотой 30-40 Гц, тогда как тельца Пачини - на стимулы с частотой 250 Гц. К медленно адаптирующимся кожным механорецепторам относятся диски Меркеля, имеющие точечные рецептивные поля, и тельца Руффини, активизирующиеся при растяжении кожи. Все эти рецепторы имеют миелинизированные аксоны, принадлежащие к группе волокон Аβ, за исключением рецепторов волосяных фолликулов, снабженных волокнами Λδ. Существуют механорецепторы (группа С) с немиелинизированными аксонами, которые реагируют на медленно двигающиеся стимулы (поглаживание).
Терморецепторы чувствительны к температурному воздействию. Выделяют холодовые и тепловые терморецепторы. Как правило, они относятся к адаптирующимся рецепторам, но могут отвечать и фазическим (кратковременным высокочастотным) разрядом на быстрые изменения температуры. В отличие от других рецепторов терморецепторы обладают спонтанной импульсацией в нормальных физиологических условиях и проявляют активность в широком диапазоне температур. При умеренной температуре кожи (примерно 35 °С) активны как холодовые, так и тепловые рецепторы, генерирующие высокочастотные импульсные залпы, позволяющие ЦНС получить дифференцированную информацию о колебаниях активности рецепторов, подвергающихся воздействию повышенных или пониженных температур. При согревании кожи до температуры выше 37 °С импульсация от холодовых рецепторов прекращается, при охлаждении кожи ниже 35 °С прекращается импульсация от тепловых рецепторов, при этом активируется еще один класс холодовых рецепторов - высокопороговые. Важно отметить, что при повышении температуры выше 45 °С, т.е. при достижении болевого (повреждающего) уровня, терморецепторы утрачивают активность и не сигнализируют организму об опасности развития ожога и боли. При снижении температуры кожи до определенного уровня. Большинство холодовых рецепторов снабжается волокнами Аδ, а большинство тепловых рецепторов - волокнами С. Болевые рецепторы (ноцицепторы) реагируют на угрожающие организму стимулы (рис. 3.2). Существуют два основных типа кожных ноцицепторов: Аδ-механоноцицепторы и полимодальные С-ноцицепторы. Механоноцицепторы иннервируются тонкими миелинизированными, а полимодальные С-ноцицепторы - немиелинизированными С-волокнами. Аδ-ме-
1 - нисходящий путь в составе заднебокового канатика; 2 - толстые миелинизированные афферентные волокна; 3 - Аб/С волокна; 4 - воспаление; 5 - высвобождение цитокинов; 6 - активация тучной клетки; 7 - высвобождение гистамина; 8 - Аб/С волокна; 9 - субстанция Р; 10 - симпатическая иннервация кожи и кровеносных сосудов; 11- расширение сосудов и отек; 12 - механическое повреждение; 13 - высвобождение брадикинина, простагландинов ханоноцицепторы отвечают на сильное механическое раздражение кожи, например укол иглой или щипок пинцетом. Обычно они не реагируют на термические и болевые стимулы, если не были предварительно сенситизированы. Полимодальные С-ноцицепторы реагируют на болевые стимулы разного вида - механические, термические и химические. Повышение чувствительности афферентных волокон ноцицепторов называется их сенситизацией. Как правило, сенситизация наступает после ответа рецепторов на повреждающий стимул. Сенситизированные ноцицепторы интенсивнее реагируют на повторный стимул и вызывают более сильную боль в ответ на стимул той же интенсивности в результате снижения болевого порога. Ноцицепторы способны генерировать фоновый разряд, что сопровождается появлением спонтанной боли. К сенситизации приводят повреждение или воспаление тканей вблизи от болевых нервных окончаний, которое сопровождается высвобождением тканевых (гистамин, серотонин, простагландины, лейкотриены, ионы К+ и пр.) альгогенов и действием циркулирующих медиаторов отека и воспаления (брадикинин, каллидин) (см. рис. 3.2). При активации ноцицептора из немиелинизированных С- афферентов могут высвобождаться регуляторные пептиды (субстанция Р, нейрокинин А, белок, кодируемый геном кальцитонина) (рис. 3.3). Образующийся при этом нервный импульс, помимо ортодромного, может иметь и антидромное распространение (аксон-рефлекс), в результате чего в ткани высвобождаются регуляторные нейропептиды, увеличивающие тканевую проницаемость и способствующие повышению локальной концентрации альгогенов. Эти пептиды вызывают расширение сосудов и повышение
Схема аксон-рефлекса. (Б). 7 - ноцицептивное афферентное волокно (группа IV); 8 - кожа; 9 - нервный импульс; 10 - нервное окончание; 11 - вещество; 12 - кровеносный сосуд; 13 - вазодилатация, усиление кровотока, увеличение проницаемости проницаемости капилляров, таким образом усиливая действие других веществ, которые выходят из поврежденных клеток, а также из тромбоцитов, тучных клеток и лейкоцитов, мигрирующих в патологический очаг. Развивающееся в итоге воспаление сопровождается покраснением и повышением температуры вследствие усиленного кровотока, отеком, болью и повышенной чувствительностью, обусловленной сенситизацией ноцицепторов. Рецепторы мышц, суставов и внутренних органов. Ноцицепторы реагируют при надавливании на мышцу, при высвобождении некоторых метаболитов, особенно во время ишемии. Ноцицепторы мышц иннервируются аксонами среднего диаметра и тонкими миелинизированными аксонами (группы II и III) или немиелинизированными афферентами (группа IV). Рецепторы еще одной группы, снабжаемые тонкими афферентными волокнами, классифицируются как эргорецепторы, поскольку они создают ощущение работы мышц. Скелетные мышцы содержат рецепторы нескольких типов (механорецепторы, ноцицепторы, реже термо- и хеморецепторы). Наиболее важную роль играют рецепторы растяжения мышц, к которым относятся мышечные веретена и сухожильные тельца Гольджи-Маццони. Эти рецепторы необходимы для восприятия позы тела (проприоцепции). Кроме того, они играют важную роль в регуляции движений. Мышечные веретена присутствуют в большинстве скелетных мышц, особенно в большом количестве в мышцах, требующих тонкой регуляции движений (например, в мелких мышцах кисти), и в крупных мышцах, содержащих медленные фазические волокна (волокна типа I). Диаметр мышечного веретена равен примерно 100 мкм, длина - до 10 мм. Мышечное веретено состоит из пучка модифицированных мышечных волокон, иннервируемых сенсорными и двигательными аксонами (рис. 3.4). Зона иннервации мышечного веретена заключена в соединительнотканную капсулу. Мышечное веретено свободно располагается в мышце, ориентируясь параллельно мышечным волокнам. Его дистальный конец прикреплен к соединительнотканной сети внутри мышцы - эндомизию. Мышечное веретено содержит модифицированные мышечные волокна, называемые интрафузальными, в отличие от обычных - экстрафузальных. Интрафузальные волокна гораздо тоньше, чем экстрафузальные, и слишком слабы, чтобы участвовать в сокращении мышцы. Различают два типа интрафузальных мышечных волокон: с ядерной сумкой и с ядерной цепочкой (рис. 3.5). Волокна с ядерной сумкой крупнее, чем волокна с ядерной цепочкой, и их ядра плотно упакованы в средней части волокна, тогда как в волокнах с ядерной цепочкой все ядра расположены в один ряд.
1 - сухожильный орган Гольджи; 2 - капсула; 3 - соединительная капсула; 4 - мышечное веретено; 5 - первичное чувствительное окончание (волокна Iа типа); 6 - вторичное чувствительное окончание (волокна II типа); 7 - 7-эфферентное двигательное волокно; 8 - α-эфферентное двигательное волокно к экстрафузальным мышечным волокнам; 9 - экстрафузальные мышечные волокна; 10 - интрафузальные мышечные волокна; 11 - чувствительное волокно; 12 - сухожилие Сенсорная иннервация мышечных веретен осуществляется одним афферентным аксоном группы Ia и несколькими афферентными аксонами группы II (см. рис. 3.5). Афференты Ia являются сенсорными аксонами наибольшего диаметра со скоростью проведения от 72 до 120 м/с; они образуют первичное окончание, спирально обвивая каждое интрафузальное волокно. Первичные окончания располагаются на интрафузальных волокнах обоих типов (с ядерной сумкой и с ядерной цепочкой), что важно для деятельности этих рецепторов. Аксоны группы II имеют промежуточный диаметр и проводят импульсы со скоростью от 36 до 72 м/с; они образуют вторичные окончания только на волокнах с ядерной цепочкой. Двигательную иннервацию мышечных веретен обеспечивают два типа γ-эфферентных аксонов (см. рис. 3.5). Динамические 7-эфференты оканчиваются на каждом волокне с ядерной сумкой, статические 7-эфференты - на волокнах с ядерной цепочкой. Мышечное веретено реагирует на растяжение мышцы. Сокращение экстрафузальных мышечных волокон заставляет мышечное веретено укорачиваться, поскольку оно лежит параллельно экстрафузальным волокнам. Активность афферентов мышечных веретен зависит от механического растяжения афферентных окончаний на интрафузальных волокнах. При сокращении экстрафузальных волокон мышечное волокно укорачивается, расстояние между витками афферентного нервного окончания уменьшается и частота разряда в афферентном аксоне падает. Нао-
1 - Динамическое 7-эфферентное волокно; 2 - статическое 7-эфферент- ное волокно; 3 - афферент группы Iа; 4 - афферент групп II; 5 - пластинчатое 7-окончание; 6 - интрафузальное мышечное волокно с ядерной сумкой; 7 - интрафузальное мышечное волокно с ядерной цепочкой; 8 - стелющееся 7-окончание; 9 - первичное окончание; 10 - вторичное окончание борот, когда вся мышца подвергается растяжению, мышечное веретено тоже удлиняется (так как его концы прикреплены к соединительнотканной сети внутри мышцы) и растягивание афферентного окончания повышает частоту его импульсного разряда. Такая реакция называется статическим ответом афферентов мышечного веретена. Первичные и вторичные афферентные окончания отвечают на растяжение по-разному. Первичные окончания чувствительны и к степени растяжения, и к его скорости, а вторичные окончания реагируют преимущественно на величину растяжения. Эти различия определяют активность окончаний двух типов. Частота разряда первичного окончания достигает максимума во время растяжения мышцы, а при расслаблении растянутой мышцы разряд прекращается. Реакция такого типа называется динамическим ответом афферентных аксонов группы Iа. Постукивание по мышце (либо по ее сухожилию) или синусоидальное растяжение более эффективно вызывает разряд в первичном афференте, чем во вторичном. В скелетных мышцах есть еще один тип рецепторов растяжения - сухожильные тельца Гольджи-Маццони, образованные толстыми миелиновыми волокнами, «намотанными» вокруг групп коллагеновых сухожильных волокон, окруженных соединительнотканной капсулой (афференты группы Ib). Рецептор имеет диаметр около 100 мкм и длину примерно 1 мм, скорость проведения импульса такая же, как и у афферентов группы Iа. Тельца Гольджи-Маццони соединены с мышцей последовательно, в отличие от мышечных веретен, расположенных параллельно экстрафузальным волокнам, что позволяет им активироваться как при сокращении, так и при растяжении мышцы. Сокращение мышцы - более эффективный раздражитель, чем растяжение, поскольку стимулом для рецептора является сила, развиваемая сухожилием, в котором он находится. Тельца Гольджи-Маццони информируют о развиваемой силе мышечного сокращения, а мышечное веретено - о длине мышцы и скорости ее изменения. Ткани суставов снабжены рецепторами, медленно и быстро адаптирующимися механорецепторами и ноцицепторами. Быстро адаптирующиеся механорецепторы - тельца Пачини - отвечают на кратковременное механическое раздражение, в том числе вибрацию. Медленно адаптирующиеся рецепторы - тельца Руффини - реагируют на смещения элементов суставов в крайние положения. Они сигнализируют о давлении на сустав или о его вращении. Механорецепторы суставов иннервируются афферентами среднего диаметра (группа II). Суставные ноцицепторы активируются при чрезмерном разгибании или сгибании в суставе, но остаются неактивными при движениях в физиологическом объеме. Если вследствие воспаления сустава ноцицепторы сенситизировались, они реагируют на слабые движения или незначительное давление, которые в нормальных условиях не вызывают их ответа. Ноцицепторы суставов иннервируются тонкими миелинизированными (группа III) или немиелинизированными (группа IV) первичными афферентами. Во внутренних органах относительно немного рецепторов. Есть висцеральные рецепторы (интероцепторы), которые участвуют в обычных рефлекторных актах, не вызывая сенсорного восприятия. Некоторые висцеральные механорецепторы опосредуют чувство наполнения органа, а висцеральные ноцицепторы сигнализируют о висцеральной боли. В брыжейке и в оболочке поджелудочной железы присутствуют тельца Пачини, подающие сигналы о кратковременных механических стимулах. В некоторых внутренних органах имеются специфичные ноцицепторы. Вероятно, некоторые висцеральные рецепторы включаются только при сенситизации, обусловленной повреждением ткани. Волокна, исходящие из нервно-мышечных веретен и имеющие толстую оболочку, занимают самую медиальную часть заднего (чувствительного) корешка. Среднюю часть корешка занимают волокна, исходящие из инкапсулированных рецепторов. Наиболее латерально расположены мало миелинизированные волокна, проводящие болевые и температурные импульсы. Только некоторые импульсы, приходящие из мышц, суставов, фасций и других тканей, достигают уровня коры большого мозга и подвергаются осознанному анализу; большинство импульсов участвует в реализации автоматического контроля двигательной активности. Проходя в спинной мозг через задние корешки, отдельные волокна разделяются на многочисленные коллатерали, которые обеспечивают синаптические связи с другими нейронами спинного мозга. Все афферентные волокна при прохождении через задние корешки лишаются миелинового покрытия и идут в различных трактах в зависимости от их чувствительной модальности. Наиболее распространена следующая классификация чувствительности: • поверхностная (экстероцептивная) - болевая, температурная и тактильная чувствительность; • глубокая (проприоцептивная) - мышечно-суставная, вибрационная чувствительность, чувство давления, массы тела, определение направления движения кожной складки (кинестезия); • сложные формы чувствительности: чувство локализации укола, прикосновения, узнавание написанных на коже знаков и букв (двухмерно-пространственное чувство), различение уколов, наносимых одновременно на близком расстоянии циркулем Вебера (дискриминационная чувствительность), стереогноз; • ощущение, обусловленное раздражением рецепторов внутренних органов (интероцептивная чувствительность). С точки зрения филогенетического развития различают протопатическую и эпикритическую чувствительность. Протопатическая чувствительность более древняя, у нее ограниченные возможности дифференциации раздражений по их модальности, интенсивности и локализации. Эпикритическая чувствительность - филогенетически новый вид, обеспечивающий возможность количественной и качественной оценки раздражений по модальности, интенсивности, локализации. Экстероцептивными являются ощущения, формирующиеся в чувствительных образованиях кожи или слизистых оболочек в ответ на внешние воздействия или изменения окружающей среды. Их также называют поверхностными, или кожными, и исходящими из слизистых оболочек видами чувствительности. Существуют три разновидности экстероцептивной чувствительности: болевая, температурная (холодовая и тепловая) и тактильная (ощущение легкого прикосновения). Проприоцептивная чувствительность исходит из глубоких тканей: мышц, связок, сухожилий, суставов. Термин «сложная чувствительность» применяется для описания тех вариантов чувствительности, осуществление которых требует присоединения кортикального компонента для достижения чувства окончательного восприятия. При этом ведущей является функция восприятия и различения по сравнению с простым ощущением в ответ на стимуляцию первичных чувствительных окончаний. Способность к восприятию и пониманию формы и характера предметов путем прикосновения и их ощупывания называется стереогнозом. Отдельным видам чувствительности соответствуют различные проводящие пути (рис. 3.6). Пути проведения поверхностной чувствительности Первым нейроном, проводящим импульсы болевой и температурной чувствительности, являются псевдоуниполярные нейроны спинномозговых узлов (рис. 3.7), периферические ветви которых (дендриты) представляют собой тонкие миелиновые и безмиелиновые волокна, направляющиеся к рецепторам соответствующего участка кожи (дерматому). Центральные ветви этих клеток (аксоны) входят в спинной мозг через латеральную зону задних корешков. В спинном мозге они разделяются на короткие восходящие и нисходящие коллатерали, которые через 1-2 сегмента образуют синаптические контакты с интернейронами желатинозной
субстанции. Сложно организованные клеточные группы спинного мозга обеспечивают первичный анализ сенсорной информации, модуляцию сигналов и их проведение к вышележащим отделам нервной системы. В задних рогах расположены тела вторых нейронов, восходящие аксоны которых, направляясь косо вверх, через переднюю серую спайку переходят на противоположную сторону спинного. Располагаясь в блоковых столбах, они формируют латеральный спинно-таламический путь. Волокна, идущие в его составе, имеют соматотопическое распространение: волокна от нижних сегментов спинного мозга (обеспечивают иннервацию ног, нижних отделов туловища) расположены более латерально, волокна от верхних сегментов - более медиально (эксцентрическое расположение длинных проводников). Латеральный спинно-таламический путь оканчивается в вентролатеральном ядре таламуса (третий нейрон). Аксоны клеток этого ядра направляются через заднюю треть задней ножки внутренней капсулы и лучистый
A - пути поверхностной чувствительности: 1 - рецептор; 2 - спинномозговой (чувствительный) узел (первый нейрон); 3 - зона Лиссауэра; 4 - задний рог; 5 - боковой канатик; 6 - латеральный спинно-таламический путь (второй нейрон). 7 - медиальная петля; 8 - таламус; 9 - третий нейрон; 10 - кора большого мозга; Б - пути глубокой чувствительности: 1 - рецептор; 2 - спинномозговой (чувствительный) узел (первый нейрон); 3 - задний канатик; 4 - передний спинно-таламический путь (второй нейрон тактильной чувствительности); 5 - внутренние дугообразные волокна; 6 - тонкое и клиновидное ядра (второй нейрон глубокой чувствительности); 7 - медиальная петля; 8 - таламус; 9 - третий нейрон; 10 - кора большого мозга венец к коре постцентральной извилины (поля 1, 2 и 3). В постцентральной извилине имеется соматотопическое распределение, аналогичное наблюдающемуся в прецентральной извилине: в верхних отделах извилины находятся корковые центры чувствительности для нижней конечности, в средних отделах - для туловища и верхних конечностей, в нижних отделах - для лица и головы. Части тела, обладающие более тонкой способностью к дифференциации сенсорных стимулов, имеют большее представительство в постцентральной извилине («перевернутый вниз головой человек с непропорциональными по размеру частями тела»; рис. 3.8). Ход волокон, проводящих болевую чувствительность от внутренних органов, такой же, как и волокон соматической болевой чувствительности. Осуществляет проведение передний спинно-таламический путь. Первым нейроном путей тактильной чувствительности также являются клет-
1 - глотка; 2 - язык; 3 - зубы, десны, челюсть; 4 - нижняя губа; 5 - верхняя губа; 6 - лицо; 7 - нос; 8 - глаза; 9 - I палец кисти; 10 - II палец кисти; 11 - III и IV пальцы кисти; 12 - V палец кисти; 13 - кисть; 14 - запястье; 15 - предплечье; 16 - локоть; 17 - плечо; 18 - голова; 19 - шея; 20 - туловище; 21 - бедро; 22 - голень; 23 - стопа; 24 - пальцы стопы; 25 - половые органы ки спинномозгового узла. Их умеренно толстые миелинизированные периферические волокна оканчиваются в соответствующих дерматомах, а аксоны через задний корешок вступают в задний рог спинного мозга, где переключаются на второй нейрон. Параллельно волокнам, проводящим болевую и температурную чувствительность, через белую спайку они переходят на противоположную сторону, но в отличие от них к таламусу поднимаются в составе переднего спинноталамического пути, проходящего в переднем канатике. Объединяясь в стволе мозга с латеральным спинноталамическим путем в медиальную петлю, также заканчивается в вентролатеральном ядре таламуса (третий нейрон). Его аксоны через внутреннюю капсулу с лучистый венец проводят импульсы в постцентральную извилину. Пути проведения глубокой чувствительности Проприоцептивные импульсы исходят из рецепторов мышц, сухожилий, фасций, капсул суставов, глубокой соединительной ткани и кожи (см. рис. 3.7) и проводятся в спинной мозг по отросткам псевдоуниполярных нейронов спинномозговых узлов (первый нейрон). Отдав коллатерали к нейронам задних и передних рогов серого вещества, основная часть аксонов первого нейрона входит в задний канатик. Часть аксонов идет вниз, другие - вверх в составе медиального тонкого пучка (Голля) и латерального клиновидного пучка (Бурдаха) и оканчиваются в собственных ядрах: тонком и клиновидном, расположенных на дорсальной стороне покрышки нижней части продолговатого мозга (второй нейрон). Волокна, поднимающиеся в составе задних канатиков, расположены в соматотопическом порядке. Те из них, которые проводят импульсы от области промежности, ног, нижней половины туловища, идут в тонком пучке, прилежащем к задней срединной борозде. Другие, проводящие импульсы от груди, рук и шеи, проходят в составе клиновидного пучка, причем волокна от шеи расположены наиболее латерально. Аксоны вторых нейронов, поднимаясь к таламусу, формируют бульботаламический путь. Он проходит сначала кпереди над перекрестом пирамидных путей, затем в составе медиальной петли переходит на противоположную сторону и поднимается кзади от пирамид и медиально от нижних олив через продолговатый мозг, мост и средний мозг к вентролатеральному ядру таламуса, где лежат третьи нейроны. Их аксоны нервных клеток этого ядра образуют таламокортикальный путь, который проходит через заднюю треть задней ножки внутренней капсулы и лучистый венец белого вещества головного мозга и оканчивается в постцентральной извилине (поля 1, 2, 3) и верхней теменной дольке (поля 5 и 7). Соматотопическая организация сохраняется на протяжении всего хода волокон к таламусу и коре (см. рис. 3.8). Не все афферентные импульсы передаются таламусом в чувствительную область коры - часть из них оканчивается в двигательной области коры в прецентральной извилине. Моторные и сенсорные кортикальные поля до определенной степени перекрываются, поэтому можно говорить о центральных извилинах как о сенсомоторной области. Чувствительные сигналы здесь могут быть немедленно преобразованы в двигательные реакции, что обеспечивает существование сенсомоторных кругов обратной связи. Пирамидные волокна этих коротких кругов обычно оканчиваются прямо на клетках передних рогов спинного мозга без вставочных нейронов. Поступившие в ЦНС афферентные импульсы распространяются прежде всего по специфическим для данной сенсорной модальности проекционным путям в корковые отделы анализатора. Вместе с тем на уровне среднего мозга от волокон специфических сенсорных путей отходят коллатерали, по которым возбуждение иррадиирует в ретикулярную формацию, неспецифические ядра таламуса и гипоталамуса, структуры лимбической системы и мозжечок. Мощный поток импульсов от мышц, сухожилий, суставов и глубоких тканей идет к мозжечку по спинно-мозжечковым путям. В заднем роге спинного мозга расположены клетки, дающие начало восходящим спинно-покрышечному, спинно-ретикулярному, спинооливному, спинопреддверному путям. Тем самым обеспечивается возможность регуляции высшими отделами ЦНС состояния рецепторной и проводниковой части анализатора. Это позволяет организму из многих раздражителей активно отбирать наиболее значимую в данный момент информацию. Методика исследования чувствительности Для выявления сенсорных феноменов, определения их характера и выраженности необходимо выяснить, беспокоят ли пациента боли, имеется ли утрата чувствительности, есть ли ощущение онемения в какой-либо части тела, испытывает ли он ощущение жжения, давления, растяжения, пощипывания, «ползания мурашек» и др. Как правило, исследование чувствительной сферы рекомендуется проводить в начале обследования. Это простое, на первый взгляд, исследование нужно осуществлять осторожно и тщательно. Следует избегать подсказок в описании ощущений, их интенсивности, эмоциональной окраски. Оценка результатов основывается на субъективных ответах больного, но часто объективные симптомы (вздрагивание больного, болевая гримаса, отдергивание конечности) помогают уточнить зону изменений чувствительности. Для подтверждения результатов чувствительность необходимо исследовать дважды. Если больной не отмечает расстройств чувствительности, врач может проверить чувствительность в соответствии с зонами невральной и сегментарной иннервации на коже лица, тела, конечностей. При обнаружении нарушений чувствительности нужно провести тщательное обследование для определения их характера и уточнения границ. Выявленные изменения отмечаются карандашом на коже больного и обозначаются на схеме. Исследование поверхностной чувствительности. Для проверки болевой чувствительности пользуются обычной иглой, глаза больного при исследовании должны быть закрыты. Покалывание следует производить то острием, то головкой иглы, чтобы обследуемый определил характер раздражителя («остро» или «тупо»), и двигаться от зон с меньшей чувствительностью к зонам с большей. Если уколы наносить слишком близко и часто, возможна суммация ощущений; если проведение замедлено, ответ больного будет соответствовать предыдущему раздражению. Температурная чувствительность проверяется с помощью пробирок с холодной (5-10 °С) и горячей (40-45 °С) водой. Больного просят отвечать «горячо» или «холодно». Обе разновидности температурных ощущений выпадают одновременно, хотя иногда одна может быть частично сохранена. (Обычно область нарушений тепловой чувствительности шире, чем холодовой.) Для оценки тактильной чувствительности к коже прикасаются кисточкой, клочком ваты, пером, кончиками пальцев. Тактильная чувствительность оценивается вместе с болевой (прикосновение попеременно острием и головкой иглы). Раздражение следует наносить легко, не производя давления на подкожные ткани. Исследование глубокой чувствительности. Для исследования суставномышечного чувства полностью расслабленный палец пациента исследующий должен охватить с боковых поверхностей с минимальным давлением и пассивно двигать его (рис. 3.9). Исследуемый палец должен быть отделен от других пальцев. Больному при этом не разрешается производить какие-либо активные движения пальцами. Если чувство движения или положения в пальцах утрачено, нужно исследовать более проксимальные части тела. В норме обследуемый должен определить движение в межфаланговых суставах с размахом в 1-2°. Сначала нарушается рас-
1 - пальцы правой руки исследующего; 2 - пальцы левой руки исследующего, фиксирующие межфаланговый сустав больного познавание положения пальцев, затем утрачивается ощущение движения. В дальнейшем эти ощущения могут быть утрачены во всей конечности. Мышечно-суставное чувство можно проверить и другим приемом: руке или пальцам больного исследующий придает определенное положение, причем глаза больного должны быть закрыты; затем больного просят описать положение руки или имитировать это положение другой рукой. Следующий прием: руки вытянуты вперед; при нарушении мышечно-суставного чувства пораженная рука совершает волнообразные движения, падает или не доводится до уровня другой руки. Для выявления сенсорной атаксии проводят пальценосовую и пяточно-коленную пробы, пробу Ромберга, оценивают походку. Вибрационная чувствительность проверяется с помощью камертона (128 или 256 Гц), установленного на костном выступе. Обращают внимание на интенсивность и длительность ощущения вибрации. Камертон приводят в состояние максимальной вибрации и устанавливают на первом пальце либо на медиальной или латеральной лодыжке и держат, пока больной ощущает вибрацию. Затем камертон следует установить на запястье, грудину или ключицу и уточнить, ощущает ли больной вибрацию. Можно также сравнить ощущение вибрации больного и исследующего. Для исследования чувства давления нажимают на подкожные ткани: мышцы, сухожилия, нервные стволы. Можно пользоваться тупым предметом, а также сжимать ткани между пальцами. Уточняют восприятие давления и его локализацию. Для количественной оценки используют эстезиометр, в котором дифференцировка локального давления определяется в граммах. Для выявления чувства массы больному предлагают определить разницу в массе двух одинаковых по форме и величине предметов, положенных на ладони. Исследование кинестетической чувствительности (определение направления кожной складки): больной должен с закрытыми глазами определить, в каком направлении исследующий двигает кожную складку на туловище, руке, ноге - вверх или вниз. Исследование сложной чувствительности. Чувство локализации уколов и прикосновения к коже определяют у больного с закрытыми глазами. Дискриминационную чувствительность (способность различать два наносимых одновременно раздражения кожи) исследуют циркулем Вебера или калиброванным двухмерным эстезиометром. Больной с закрытыми глазами должен определить минимальное расстояние между двумя точками воздействия. Это расстояние разное на разных частях тела: 1 мм на кончике языка, 2-4 мм на ладонной поверхности кончиков пальцев, 4-6 мм на тыльной поверхности пальцев, 8-12 мм на ладони, 20-30 мм на тыльной стороне кисти. Большее расстояние наблюдается на предплечье, плече, теле, голени и бедре. Двумерно-пространственное чувство - узнавание знаков, написанных на коже. Обследуемый с закрытыми глазами должен определить буквы и цифры, которые исследующий пишет на коже. Стереогноз - узнавание предмета на ощупь. Больной с закрытыми глазами ощупывает положенный в руку предмет и определяет его форму, величину, консистенцию. Расстройства чувствительности Болевое ощущение является наиболее частым симптомом заболевания и причиной обращения к врачу. Боль при заболеваниях внутренних органов возникает вследствие нарушения кровотока, спазма гладкой мускулатуры, растяжения стенок полых органов, воспалительных изменений в тканях. Поражение вещества мозга не сопровождается болью, она возникает при раздражении оболочек, внутричерепных сосудов. Боли могут возникать в связи с раздражением чувствительных корешков и нервных стволов, нередко они бывают проекционными, т.е. ощущаются не только в месте раздражения, но и дистальнее - в области, иннервируемой этими нервами и корешками. К проекционным также относятся фантомные боли в отсутствующих сегментах конечностей после ампутации, центральные боли, особенно мучительные при поражении таламуса. Боли могут быть иррадиирующими, т.е. распространяющимися с одной из ветвей нерва на другие, непосредственно не вовлеченные в патологический процесс. Боль может проявляться в зоне сегментарной иннервации или в отдаленном участке; в зоне, непосредственно не связанной с патологическим очагом (отраженные боли). Болевая реперкуссия реализуется при участии клеток спинномозговых узлов, серого вещества спинного мозга и мозгового ствола, вегетативной нервной системы. Реперкуссия проявляется вегетативными, чувствительными, двигательными, трофическими и другими феноменами. Отраженные болевые зоны Захарьи- на-Геда возникают при иррадиации раздражения в соответствующую зону на коже при заболеваниях внутренних органов. Зоны отраженных болей: сердце соответствует сегментам СIII-СIV и ThI-ThVI, желудок - СIII-СIV и ThVI-ThIX, кишечник - ThIX-ThXII, печень и желчный пузырь - ThVII - ThIX, почка и мочеточник - ThXI-SI, мочевой пузырь - ThXI-SIV, матка - ThIX-SIV. При невралгических болях и невритах можно обнаружить болезненность мышц и нервных стволов при их пальпации и растяжении. Пальпацию производят там, где нервы расположены близко к костям или поверхности (болевые точки). Tаковы болевые точки затылочного нерва книзу от затылочных бугров; надключичная, соответствующая плечевому сплетению, а также токи Вале по ходу седалищного нерва. Боль может возникнуть при растяжении нерва или корешка (так называемые симптомы натяжения нервных стволов при корешковом типе поражения). Симптом Ласега характерен для поражения седалищного нерва: у лежащего на спине обследуемого разогнутую в коленном суставе ногу поднимают кверху в тазобедренном суставе (первая фаза натяжения нерва болевая), затем сгибают голень (вторая фаза - исчезновение боли вследствие прекращения натяжения нерва). Симптом Мацкевича (рис. 3.10) возникает при поражении бедренного нерва: максимальное сгибание голени у больного, лежащего на животе, вызывает боль на передней поверхности бедра. При поражении этого нерва определяется и симптом Вассермана (рис. 3.11): если больному, лежащему на животе, разгибают ногу в тазобедренном суставе, то возникает боль на передней поверхности бедра. Нарушения чувствительности включают в себя гипестезию - снижение чувствительности, анестезию - отсутствие чувствительности (аналгезия - потеря болевой чувствительности), дизестезию - извращение восприятия раздражения (тактильное или тепловое раздражение ощущается как болевое и т.д.), топанестезию - отсутствие чувства локализации, термоанестезию - отсутствие температурной чувствительности, астереогноз - нарушение пространственного чувства (стереогноза), гиперестезию или гипералгезию - повышение чувствительности при нормальной интенсивности раздражителя, гиперпатию - повышение порога возбудимости (легкие раздражения не воспринимаются, при более интенсивных возникают чрезмерные, стойкие болезненные ощущения), парестезии - ощущение ползания мурашек, зуда, холода, жжения, онемения и др., возникающие спонтанно или вследствие раздражения
нервных стволов без раздражения рецепторного аппарата, каузалгию - мучительные ощущения жжения на фоне интенсивных болей при неполном перерыве некоторых крупных нервных стволов, полиэстезию - восприятие одиночного раздражения как множественного, аллоэстезию - восприятие ощущения на отдалении от нанесенного раздражения; аллохейрию - ощущение раздражения в симметричном участке на противоположной стороне, фантомные боли - ощущение отсутствующей части конечности, боли в ней. Топическая диагностика нарушений чувствительности. Синдромы нарушений чувствительности различаются в зависимости от локализации патологического процесса (рис. 3.12). Поражение периферических нервов обусловливает невральный (невритический) тип расстройства чувствительности (рис. 3.13) - боль, гипестезию или анестезию (нарушаются все виды чувствительности), болевые точки в зоне иннервации, симптомы натяжения нервных стволов. Выявляемая при повреждении данного нерва зона гипестезии обычно меньше, чем анатомическая зона его иннервации, вследствие перекрытия соседними нервами. Нервы лица и туловища обычно имеют область перекрытия по средней линии (буль- шую на туловище, чем на лице), поэтому анестезия обычно ограничена областью, не пересекающей средней линии. Отмечаются боли в области пораженного нерва (гиперпатия, гипералгезия). Боль усиливается при надавливании на нервный ствол, точки его выхода на поверхности тканей. Одновременное поражение многих периферических нервов (полинейропатический тип) проявляется болями, парестезиями, гипоили анестезией в дистальных отделах конечностей (онемение по типу «носков» и «перчаток»). Плексалгический тип (при поражении сплетения) проявляется болью, симптомами натяжения нервов, идущих из сплетения, нарушениями чувствительности в соответствующей зоне иннервации. Обычно при этом имеются и двигательные расстройства. Радикулярный тип (при поражении задних корешков) - парестезии, боль, нарушения всех видов чувствительности в соответствующих зонах
Красным цветом обозначено нарушение всех видов чувствительности, голубым - поверхностной чувствительности, зеленым - глубокой чувствительности иннервации, симптомы натяжения корешков, болезненность в паравертебральных точках, в области остистых отростков и по ходу проекции нервных стволов. Если поврежденные корешки иннервируют конечности, возможны также гипорефлексия, гипотония и гипотрофии иннервируемых мышц. Ганглионарный тип (при поражении спинномозгового узла) вызывает боль (часто пароксизмальную), гипестезию или гиперестезию в зоне иннервации нервов, исходящих из пораженного корешка, сопровождается герпетическими высыпаниями в этой области.
Передняя поверхность: 1 - глазной нерв (I ветвь тройничного нерва); 2 - верхнечелюстной нерв (II ветвь тройничного нерва); 3 - нижнечелюстной нерв (III ветвь тройничного нерва); 4 - поперечный нерв шеи; 5 - надключичные нервы (латеральные, промежуточные, медиальные); 6 - подмышечный нерв; 7 - медиальный кожный нерв плеча; 8 - задний кожный нерв плеча; 8а - межреберно-плечевой нерв; 9 - медиальный кожный нерв предплечья; 10 - латеральный кожный нерв предплечья; 11 - лучевой нерв; 12 - срединный нерв; 13 - локтевой нерв; 14 - латеральный кожный нерв бедра; 15 - передняя ветвь запирательного нерва; 16 - передние кожные ветви бедренного нерва; 17 - общий малоберцовый нерв; 18 - подкожный нерв (ветвь бедренного нерва); 19 - поверхностный малоберцовый нерв; 20 - глубокий малоберцовый нерв; 21 - бедренно-половой нерв; 22 - подвздошно-паховый нерв; 23 - передняя кожная ветвь подвздошно-подчревного нерва; 24 - передние кожные ветви межреберных нервов; 25 - латеральные кожные ветви межреберных нервов Симпаталгический тип (при поражении симпатических ганглиев) вызывает каузалгию, резкие иррадиирующие боли, вазомоторно-трофические расстройства в зоне нарушенной иннервации. При поражении задних рогов и передней белой спайки спинного мозга наблюдаются сегментарные расстройства чувствительности - снижение болевой и температурной чувствительности в соответствующих дерматомах при сохранности глубокой. Дерматомы соответствуют сегментам спинного мозга, что имеет диагностическую значимость в определении уровня его поражения. На рис. 3.14-3.15 представлены границы шейной, грудной, по-
Задняя поверхность: 1 - большой затылочный нерв; 2 - малый затылочный нерв; 3 - большой ушной нерв; 4 - поперечный нерв шеи; 5 - подзатылочный нерв; 6 - латеральные надключичные нервы; 7 - медиальные кожные ветви (от задних ветвей грудных нервов); 8 - латеральные кожные ветви (от задних ветвей грудных нервов); 9 - подмышечный нерв; 9а - межреберноплечевой нерв; 10 - медиальный кожный нерв плеча; 11 - задний кожный нерв плеча; 12 - медиальный кожный нерв предплечья; 13 - задний кожный нерв предплечья; 14 - латеральный кожный нерв предплечья; 15 - лучевой нерв; 16 - срединный нерв; 17 - локтевой нерв; 18 - латеральная кожная ветвь подвздошно-подчревного нерва; 19 - латеральный кожный нерв бедра; 20 - передние кожные ветви бедренного нерва; 21 - запирательный нерв; 22 - задний кожный нерв бедра; 23 - общий малоберцовый нерв; 24 - поверхностный малоберцовый нерв; 25 - подкожный нерв; 26 - икроножный нерв; 27 - латеральный подошвенный нерв; 28 - медиальный подошвенный нерв; 29 - большеберцовый нерв
А - зоны сегментарной иннервации головы: 1 - мозговой ствол; 2 - ядро тройничного нерва. Б - зоны периферической иннервации головы: 1 - большой затылочный нерв; 2 - малый затылочный нерв; 3 - подзатылочный нерв; 4 - поперечный нерв шеи; 5 - большой ушной нерв; 6 - нижнечелюстной нерв (из тройничного нерва); 7 - верхнечелюстной нерв (из тройничного нерва); 8 - глазной нерв
ясничной и крестцовой сегментарных зон иннервации. Нарушение поверхностных видов чувствительности при сохранных глубоких (или наоборот), свойственное очаговому поражению спинного мозга, обозначается как диссоциированный тип расстройств чувствительности. При поражении задних канатиков имеется нарушение глубокой чувствительности ниже уровня поражения при сохранности поверхностной чувствительности (диссоциированный тип расстройств чувствительности), при этом наблюдается сенситивная атаксия. Движения становятся несоразмерными, неточными, при выполне- нии движений включаются мышцы, не имеющие прямого отношения к выполняемому движению. При ходьбе больной чрезмерно разгибает ноги и выбрасывает их вперед, сильно топает («штампующая походка»). При включении зрения атаксия уменьшается. Атаксия в нижних конечностях выявляется при пяточно-коленной пробе, пробе Ромберга. Поражение половины спинного мозга (синдром Броун-Секара) сопровождается снижением глубокой чувствительности и центральными двигательными расстройствами на стороне поражения и нарушением поверхностной чувствительности на противоположной. При полном поперечном поражении спинного мозга наблюдается проводниковый тип расстройства всех видов чувствительности ниже уровня поражения - параанестезия. При поражении мозгового ствола возможен альтернирующий тип расстройства чувствительности: снижение поверхностной чувствительности в противоположных очагу конечностях (в результате поражения спинноталамического пути) и сегментарная гипестезия на лице на стороне очага (вследствие поражения ядра тройничного нерва). Таламический тип расстройства чувствительности (при поражении таламуса) - гемигипестезия в противоположных очагу конечностях на фоне гиперпатии, преобладание расстройств глубокой чувствительности, «таламические» боли (жгучие, периодически усиливающиеся и плохо купируемые анальгетиками). Если поражены чувствительные пути в задней ножке внутренней капсулы, выпадают все виды чувствительности на противоположной половине тела (гемигипестезия или гемианестезия). Как правило, одновременно имеется поражение и других проводящих путей (гемипарез, центральное поражение лицевого и подъязычного нервов, гемианопсия). Корковый тип расстройства чувствительности (при поражении коры больших полушарий) проявляется парестезиями (покалывание, ползание мурашек, онемение) в половине верхней губы, языка, лица, в руке или ноге на противоположной стороне в зависимости от локализации поражения в постцентральной извилине. Парестезии могут иметь вид фокальных чувствительных эпилептических припадков (парциальные сенсорные припадки). Распознавание объектов при прикосновении (стереогноз) требует включения дополнительных ассоциативных полей коры, локализованных в теменной доле, где информация о размере, форме, физических свойствах (острота, мягкость, твердость, температура и др.) объекта интегрируется и может быть сопоставлена с теми тактильными ощущениями, которые имелись в прошлом. В связи с этим поражение нижней теменной дольки проявляется астереогнозом, т.е. утратой способности узнавать предметы при прикосновении к ним на противоположной очагу стороне. Нарушение мышечно-суставной чувствительности означает расстройство координации движений, неловкость при выполнении произвольных движений, гиперметрию. Это может проявляться в виде афферентного пареза, т.е. расстройств двигательных функций, которые обусловлены нарушением мышечно-суставного чувства при сохранной мышечной силе. Синдром афферентного пареза может быть одним из признаков поражения теменной доли. Дата добавления: 2015-12-16 | Просмотры: 205633 | Нарушение авторских прав |
 Рис. 3.1. Классификация рецепторов по скорости их адаптации и характеристикам соответствующих им рецепторных полей. (А). Распределение рецепторов, расположенных в коже, лишенной волосяного покрова. (Б). 1 - тельца Пачини; 2 - тельца Руффини; 3 - диски Меркеля; 4 - тельца Мейсснера; 5 - эпидермис; 6 - периферический нерв; 7 - дерма
Рис. 3.1. Классификация рецепторов по скорости их адаптации и характеристикам соответствующих им рецепторных полей. (А). Распределение рецепторов, расположенных в коже, лишенной волосяного покрова. (Б). 1 - тельца Пачини; 2 - тельца Руффини; 3 - диски Меркеля; 4 - тельца Мейсснера; 5 - эпидермис; 6 - периферический нерв; 7 - дерма Рис. 3.2. Схема работы ноцицептивной системы.
Рис. 3.2. Схема работы ноцицептивной системы. Рис. 3.3. Механизм сенситизации нервных ноцицептивных окончаний. (А). 1 - нервное окончание; 2 - сенситизация; 3 - синтез брадикинина; 4 - взаимодействие с γ-глобулинами крови; 5 - выход протеолитических ферментов; 6 - повреждение. Гибель клетки.
Рис. 3.3. Механизм сенситизации нервных ноцицептивных окончаний. (А). 1 - нервное окончание; 2 - сенситизация; 3 - синтез брадикинина; 4 - взаимодействие с γ-глобулинами крови; 5 - выход протеолитических ферментов; 6 - повреждение. Гибель клетки.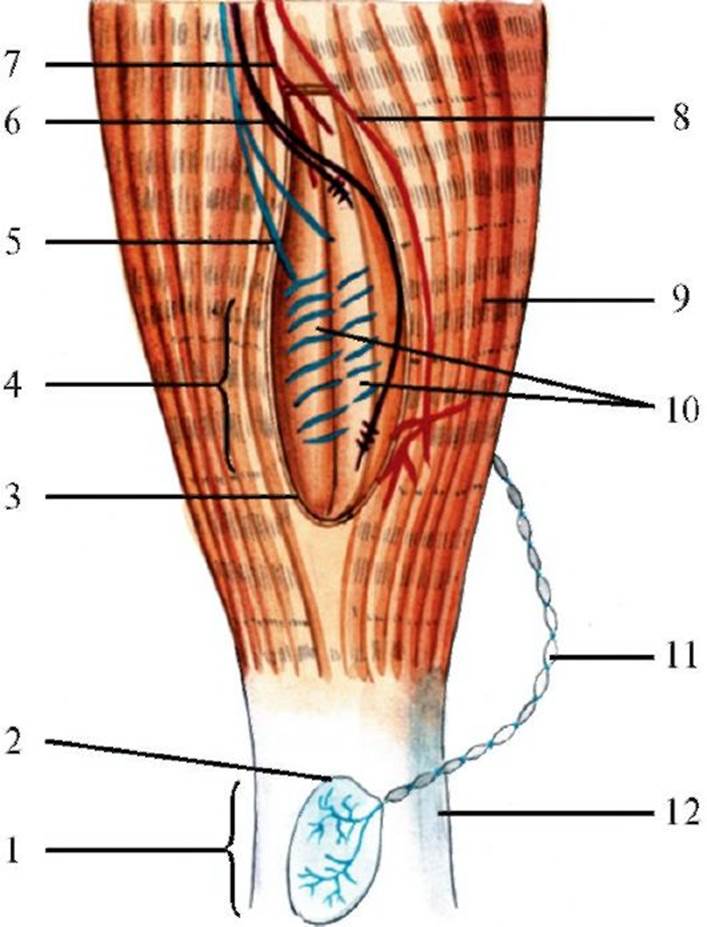 Рис. 3.4. Схема мышечного веретена.
Рис. 3.4. Схема мышечного веретена. Рис. 3.5. Типы интрафузальных мышечных волокон.
Рис. 3.5. Типы интрафузальных мышечных волокон. Рис. 3.6. Рецепторы нервных волокон заднего корешка спинного мозга. 1, 2 - большие ганглиозные клетки задних корешков, аксоны которых идут к задним канатикам, а афферентные волокна начинаются от пачиниевых телец и мышечных веретен; 3, 4 - клетки задних корешков, аксоны которых заканчиваются на клетках задних рогов спинного мозга, дающих начало спинно-таламическому и спинно-мозжечковому путям; 5 - клетки, аксоны которых заканчиваются на нейронах задних рогов спинного мозга, дающих начало переднему спинно-таламическому пути; 6 - тонкие волокна болевой чувствительности, заканчивающиеся в студенистом веществе, откуда берут начало волокна спинноталамического пути: I - медиальная часть; II - латеральная часть
Рис. 3.6. Рецепторы нервных волокон заднего корешка спинного мозга. 1, 2 - большие ганглиозные клетки задних корешков, аксоны которых идут к задним канатикам, а афферентные волокна начинаются от пачиниевых телец и мышечных веретен; 3, 4 - клетки задних корешков, аксоны которых заканчиваются на клетках задних рогов спинного мозга, дающих начало спинно-таламическому и спинно-мозжечковому путям; 5 - клетки, аксоны которых заканчиваются на нейронах задних рогов спинного мозга, дающих начало переднему спинно-таламическому пути; 6 - тонкие волокна болевой чувствительности, заканчивающиеся в студенистом веществе, откуда берут начало волокна спинноталамического пути: I - медиальная часть; II - латеральная часть Рис. 3.7. Проводящие пути чувствительности (схема).
Рис. 3.7. Проводящие пути чувствительности (схема).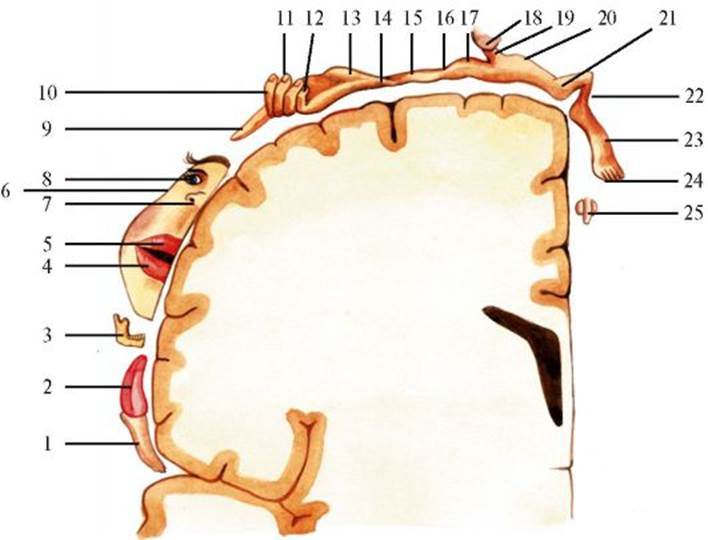 Рис. 3.8. Представительство чувствительных функций в задней центральной извилине (схема).
Рис. 3.8. Представительство чувствительных функций в задней центральной извилине (схема). Рис. 3.9. Исследование мышечно-суставного чувства.
Рис. 3.9. Исследование мышечно-суставного чувства. Рис. 3.10. Симптом Мацкевича
Рис. 3.10. Симптом Мацкевича Рис. 3.11. Симптом Вассермана
Рис. 3.11. Симптом Вассермана Рис. 3.12. Чувствительные нарушения при различных уровнях поражения нервной системы (схема). 1 - полиневритический тип; 2 - поражение шейного корешка (CVI); 3 - начальные проявления интрамедуллярного поражения грудного отдела спинного мозга (ThIV-ThIX); 4 - выраженные проявления интрамедуллярного поражения грудного отдела спинного мозга (ThIV-ThIX); 5 - полное поражение сегмента ThVII; 6 - поражение левой половины спинного мозга в шейном отделе (CIV); 7 - поражение левой половины спинного мозга в грудном отделе (ThIV); 8 - поражение конского хвоста; 9 - левостороннее поражение в нижнем отделе мозгового ствола; 10 - правостороннее поражение в верхнем отделе мозгового ствола; 11 - поражение правой теменной доли.
Рис. 3.12. Чувствительные нарушения при различных уровнях поражения нервной системы (схема). 1 - полиневритический тип; 2 - поражение шейного корешка (CVI); 3 - начальные проявления интрамедуллярного поражения грудного отдела спинного мозга (ThIV-ThIX); 4 - выраженные проявления интрамедуллярного поражения грудного отдела спинного мозга (ThIV-ThIX); 5 - полное поражение сегмента ThVII; 6 - поражение левой половины спинного мозга в шейном отделе (CIV); 7 - поражение левой половины спинного мозга в грудном отделе (ThIV); 8 - поражение конского хвоста; 9 - левостороннее поражение в нижнем отделе мозгового ствола; 10 - правостороннее поражение в верхнем отделе мозгового ствола; 11 - поражение правой теменной доли. Рис. 3.13а. Распределение кожной чувствительности соответственно нервам и сегментам спинного мозга (схема).
Рис. 3.13а. Распределение кожной чувствительности соответственно нервам и сегментам спинного мозга (схема). Рис. 3.136. Распределение кожной чувствительности соответственно нервам и сегментам спинного мозга (схема).
Рис. 3.136. Распределение кожной чувствительности соответственно нервам и сегментам спинного мозга (схема). Рис. 3.14. Сегментарная иннервация кожи головы.
Рис. 3.14. Сегментарная иннервация кожи головы. Рис. 3.15. Сегментарная иннервация кожи туловища и конечностей
Рис. 3.15. Сегментарная иннервация кожи туловища и конечностей