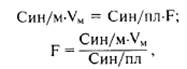|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Принцип расчета скорости клубочковой фильтрации.Скорость клубочковой фильтрации определяется объемом ультрафильтрата, образующегося в почках за единицу времени. В среднем у мужчин скорость клубочковой фильтрации составляет 125 мл/мин, у женщин — 110 мл/мин. В результате фильтрации за сутки образуется около 150—170 л первичной мочи. Эффективность фильтрации поддерживается регуляцией почечного кровотока. Из каждого объема протекающей через почки плазмы крови 0,2 объема проходит через почечный фильтр и составляет клубочковый ультрафильтрат. Большой объем ультрафильтрата является результатом: • наличия фильтрационного давления, обильного кровоснабжения почек, • обширной (до 2 м2) фильтрационной поверхности капилляров клубочков. Расчет скорости клубочковой фильтрации: Скорость клубочковой фильтрации измеряется объемом фильтрата, образующегося в почках за единицу времени. Для определения фильтрации используют диагностическое вещество инулин (полисахарид фруктозы), который вводят в кровоток: инулин попадает в мочу только путем клубочковой фильтрации, не реабсорбируется и не секретируется, а также не претерпевает метаболических превращений в канальцах. Оно беспрепятственно проходит почечный фильтр, не адсорбируется белками и содержится в фильтрате в той же концентрации, что и в плазме крови. Количество инулина, профильтровывающееся за единицу времени, равно количеству этого вещества, удаленного с мочой:
где F — объем фильтрата за 1 мин; Син/м — концентрация инулина в конечной моче; VM — объем конечной мочи за 1 мин. Из расчета видно, что объем почечной фильтрации равен клиренсу по инулину.
#90.
#91. Механизмы регуляции процессов мочеобразования. Деятельность почек находится под постоянным контролем нервных и гуморальных влияний и зависит от почечных и внепочечных факторов регуляции. Фильтрация. К почечным факторам, определяющим фильтрацию, относятся: 1. количество функционирующих клубочков и проницаемость их канальцев; 2.диаметр приносящего и выносящего сосудов; 3. давление фильтрата в капсуле. Внепочечные факторы, влияющие на фильтрацию, характеризуются: 1.общим функциональным состоянием системы кровообращения, количеством циркулирующей крови, величинами артериального давления и скорости кровотока; 2.степенью гидратации организма, осмотическим и онкотическим давлением; 3.функционированием других механизмов выделения воды (потовые железы). Реабсорбция. К почечным факторам, влияющим на реабсорбцию, относятся: 1. реабсорбционная способность почечного эпителия; 2.содержание пороговых и непороговых веществ в первичной моче. К внепочечным факторам, определяющим реабсорбцию, относятся: 1.эндокринная регуляция деятельности почечного эпителия со стороны желез внутренней секреции; 2. содержание пороговых и непороговых веществ в крови. Гуморальные факторы: Гормон коры надпочечников: альдостерон - задерживает натрий и воду, увеличивая их всасывание в почечных канальцах (калий при этом выводится). Гормон гипоталамуса: антидиуретический гормон (АДГ) или вазопрессин - сберегает воду путем уменьшения мочевыведения (повышает всасывание воды в почечных канальцах) Гормон щитовидной железы: кальцитонин - повышает выделение кальция. Гормон паращитовидной железы: паратгормон - снижает всасывание фосфора в канальцах и повышает его выведение. Содержание кальция сохраняется (снижается его выведение). Нервная регуляция осуществляется вегетативной нервной системой через чревные нервы. Активация симпатической нервной системы вызывает сужение сосудов почки и уменьшение диуреза. Конечная моча характеризуется следующими признаками: 1.ряд веществ, входящих в состав плазмы крови, полностью отсутствует в конечной моче: в норме это белки, аминокислоты, глюкоза; 2. некоторые вещества присутствуют в конечной моче в значительно больших концентрациях, чем в плазме: мочевина — в 65 раз, сульфаты — в 80 раз, мочевая кислота — в 12 раз; таким образом проявляется концентрирующая функция почек; 3.состав и реакция мочи непостоянны; например, при алкалозе моча становится более основной, а при ацидозе — более кислой. 4. образуется 1,5-2 л. конечной мочи за сутки. Диурез суточный - количество мочи, выделенное человеком за сутки. У здорового человека суточный диурез составляет 75 - 80% от принятой накануне жидкости. При нормальном питьевом режиме суточный диурез в среднем равен 1,5 л. При этом днем человек выделяет 2/3 и ночью 1/3 этого объема мочи.
#92. Механизм формирования жажды. Мотивация жажды: Возбуждение в нейронах супраоптических и паравентрикулярных ядер гипоталамуса гиперосмолярной кровью формирует ощущение и мотивацию жажды. Осморецепторы определяют обратную афферентацию в гипоталамические центры по блуждающим нервам от ЖКТ. При гиперосмолярном состоянии афферентация усиливается. Гидратация и дегидратация соединительной ткани приводит к тому, что из нее в кровь поступают специальные молекулы – олигопептиды, простагландины, которые несут информацию об осмотической потребности. Гиперосмолярная кровь. При гиперосмолярной крови сигнализация от осморецепторов, адресуясь в конечном счете к нейронам супраоптическич и паравентрикулярных ядер, усиливает образование в них вазопрессина. Вазопрессин, поступая в нейрогипофиз и в кровь, достигает своих органов-мишеней — восходящих частей петли нефронов и собирательных трубок. Происходит задержка воды в организме, что противодействует росту осмо-лярности. Дополнительно к этому за счет снижения секреции АКТГ и аль-достерона усиливается выделение натрия из организма. Под влиянием вазопрессина в мозге нарастает содержание ангиотензина II. Формируется мотивация жажды и на ее основе — питьевое повеление, приводящее к нормализации осмотического давления.
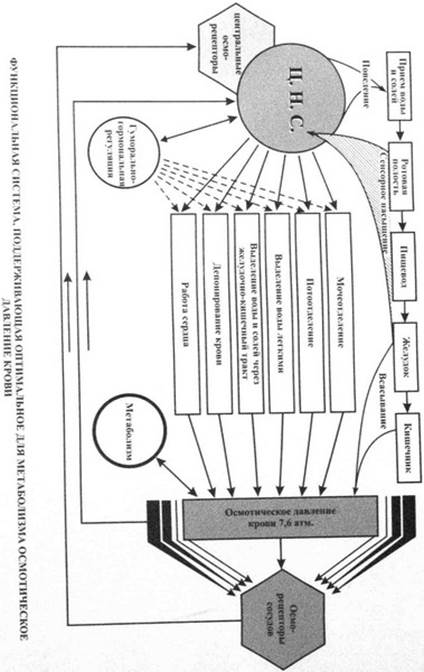
#93. Узловые механизмы функциональной системы, поддерживающей оптимальный для метаболизма уровень осмотического давления.
Оптимальный для метаболизма уровень осмотического давления определяется различным соотношением воды и ионов в организме. Оптимальным для метаболизма тканей осмотическим состоянием считается нормоосмолярное (нормоосмотическое). Преобладание солей и, наоборот, снижение воды в организме формируют гиперосмолярное (гиперосмотическое) состояние. Уменьшение содержания солей или, наоборот, при нормальном количестве солей увеличение содержания воды в организме формирует гипоосмолярное (гипоосмотическое) состояние. Предконечный результат. Функциональная система, определяющая уровень осмотического давления, имеет предконечный результат, тесно связанный с показателями осмотического давления в тканях — осмотическое давление крови. Этот предконечный результат выступает в предупредительной роли, демпфируя резкие перепады осмотического давления в тканях. Этому результату в свою очередь предшествуют другие — величина осмотического давления в разных отделах желудочно-кишечного тракта, а также специфические эмоциональные чувства жажды и солевой мотивации.
#94 Охарактеризуйте общие принципы строения и функции анализаторов. Анализатор – совокупность возбудимых структур центральной и периферической нервной системы, осуществляющих восприятие и анализ воздействий окружающей среды и воздействий, исходящих от самого организма. Классические представления Павлова об анализаторе включают в его состав три части: периферический отдел, проводниковый отдел и центральный конец. Периферический отдел анализаторов включает, как правило, рецепторы, хотя в некоторых анализаторах, например зрительном, в этот отдел могут быть включены и первичные афферентные нейроны. Периферический отдел анализатора является составной частью любого органа чувств, который, помимо рецепторов, включает специальные вспомогательные образования для наилучшего восприятия действующего раздражителя. Например, глаз как орган зрения, помимо сетчатки (фоторецепторы), включает глазное яблоко, его мышцы, веки и др. Проводниковый отдел анализаторов включает не только нервные волокна, непосредственно отходящие от рецепторов, но и все афферентные нейроны, обеспечивающие первичный анализ и передачу возбуждений в центральный отдел анализатора. Возникающие в рецепторах импульсы возбуждения распространяются по проводящим путям в виде электрических потенциалов. Во всех нервных волокнах потенциалы являются однотипными по внешнему виду, но в потоке импульсов возбуждения в их своеобразном рисунке — паттерне — закодирована специфическая информация о параметрах действующего раздражителя. Анализ этой информации начинается как на уровне первичных афферентных нервных клеток, так и в последующих спинальных, стволовых и подкорковых ядрах. Центральный отдел анализаторов. Различные проводящие афферентные пути через возбуждение соответствующих подкорковых структур в конечном счете приносят импульсы возбуждения в соответствующие области коры большого мозга, которые считаются высшим центральным конечным звеном любого анализатора. Вместе со специфическим афферентным возбуждением в кору поступает и неспецифическое восходящее возбуждение, которое формируется на уровне подкорковых активирующих структур мозга — ретикулярной формации, гипоталамуса и др. Передача импульсов от рецепторов по проводящим путям к коре большого мозга осуществляется по цепям нейронов в различных ядрах, расположенных на разных уровнях ЦНС. За счет конвергенции и дивергенции возбуждений в нейронных цепях в этих нервных центрах осуществляются передача и обработка информации. Роль анализаторов в деятельности функциональных систем: Физиологические особенности каждого анализатора в отдельности определяются его специфическими структурами передачи возбуждений от рецепторов в ЦНС, участием в системных процессах целого организма. Адекватное поведение живых организмов в окружающей среде не является пассивным отражением воздействующих раздражителей. В большей степени организм настойчиво ищет потребные раздражители и активно к ним стремится, избирательно настраивая по отношению к ним свои анализаторы. Активное стремление субъектов к раздражителям внешней среды определяется прежде всего их исходными доминирующими потребностями и пропускной способностью к передаче информации соответствующего анализатора. У человека наибольшей пропускной способностью обладает зрительный анализатор, который в единицу времени передает в ЦНСболее 70 % информации; 25—28 % информации доставляет в ЦНС слуховой анализатор и 2—5 % информации — остальные анализаторы.
#95 Рассмотрите важнейшие физиологические св-ва рецепторов. Дайте классификацию рецепторов. Рецепторы участвуют впроцессе восприятия и трансформации механической, термической, электромагнитной и химической энергии в нервный сигнал или сложную последовательность мембранных и цитоплазматических процессов. Существуют различные классификации рецепторов, основанные на их физиологических характеристиках. Психофизиологическое состояние, связанное с модальностью ощущения, в соответствии с которым выделяют зрительные, слуховые, осязательные, обонятельные, вкусовые, холодовые, тепловые, болевые рецепторы. Локализация. Большинство сенсорных рецепторов воспринимает раздражения из окружающей среды, т.е. являются внешними, или экстерорецепторами. К ним относятся фоторецепторы, слуховые, тактильные, температурные и хеморецепторы, расположенные на поверхности тела и в начальных отделах пищеварительного тракта и дыхательных путей. Информация о состоянии внутренней среды организма воспринимается интерорецепторами внутренних органов, сосудов, опорно-двигательного аппарата — мышц, сухожилий, костей, суставов. Структура рецепторов. Рецепторы могут быть представлены свободными нервными окончаниями; окончаниями, покрытыми особой капсулой (инкапсулированные; иметь вид палочек, колбочек, ветвей, волосков. Некоторые рецепторы объединяются в сложно организованные множества — сетчатку глаза, кортиев орган внутреннего уха и др. В результате рецепции действующего на организм раздражения и поступающей в мозг на ее основе сигнализации формируется субъективно переживаемое ощущение, являющееся источником познания внешнего мира. Специализация рецепторов. Характерным свойством рецепторов является их высокая генетически детерминированная специализация к восприятию адекватного раздражителя. В соответствии с природой или характером раздражения их делят на: • тактильные рецепторы кожи; • слуховые, вестибулярные и гравитационные рецепторы внутреннего уха; • рецепторы опорно-двигательного аппарата (растяжения, суставные, мышц); • барорецепторы сердца и сосудов; • хеморецепторы обоняния, вкуса, кровеносных сосудов и тканей; • фоторецепторы сетчатки — нервные элементы, возбуждаемые электромагнитными волнами дающие ощущения ахроматического — черно-белого (палочки) и хроматического — цветового (колбочки) видения; • терморецепторы кожи, внутренних органов и ЦНС, реагирующие на изменения температуры окружающей среды и внутренней среды организма. Кроме этого, выделяют рецепторы вибрации, рецепторы волосяных фолликулов, ганглиев. Модальность. Некоторые (мономодальные) рецепторы приспособлены для восприятия лишь одного вида раздражения, например вкусовые рецепторы сладкого; другие (полимодальные) — для восприятия нескольких видов раздражителей, например ноцицепторы кожи, участвующие в формировании болевого ощущения при любом механическом, химическом, температурном повреждающем воздействии. Дистантные - воспринимают информацию от источника, расположенного на некотором расстоянии от них (зрительные, слуховые) Контактные — при непосредственном соприкосновении с раздражителем (тактильные). Чувствительность. Большинство рецепторов обладает высокой чувствительностью по отношению к адекватным раздражителям Низкопороговые — наиболее чувствительные рецепторы — расположены в коже (тактильные, или осязательные, волоски), в сетчатке глаза (палочки), в обонятельных луковицах. К высокопороговым — наименее чувствительным — относятся рецепторы сетчатки (колбочки), ответственные за хроматическое (цветовое) зрение, и ноцицепторы кожи, возбуждающиеся при механическом воздействии повреждающей интенсивности. Адаптация — изменение порога чувствительности рецептора при постоянном действии на него раздражителя. #96 Проанализируйте ф-ции вспомогательного аппарата, оптической системы и рецепторного аппарата зрительного анализатора. Уметь определять остроту зрения по таблице и интерпретировать полученные результаты. Светопреломляющие структуры глаза: роговица, радужная оболочка, хрусталик, камерная влага и стекловидное тело - обеспечивают формирование на сетчатке реального, уменьшенного и перевернутого изображения объекта внешнего мира. Радужная оболочка образует зрачок. Светопреломляющая способность хрусталика и диаметр зрачка изменяются при сокращении гладких мышц глаза. Зрачковая реакция на свет является механизмом снижения количества света, падающего на сетчатку при сильном освещении (сужение зрачка), или повышения количества света при слабом освещении за счет увеличения ширины зрачка. Физиологические механизмы опознания зрительных объектов начинаются с первичной обработки зрительной информации в сетчатке глаза, которая является периферической рецепторной структурой зрительного анализатора. Сетчатка расположена на внутренней поверхности задней сферы глазного яблока и состоит из клеток пигментного эпителия, фоторецепторов и четырех слоев, образованных различными нервными клетками. Фоторецепторы сетчатки: основными зрительными рецепторами, расположенными в сетчатке, являются палочки и колбочки. У человека рецепторный слой сетчатки состоит из 120 млн палочек и 6 млн колбочек. Колбочки воспринимают цвета и функционируют в условиях яркой освещенности объектов, в то время как палочки воспринимают световые потоки в условиях сумерек. Фоторецепторы сетчатки содержат светочувствительные пигменты, которые обесцвечиваются при действии света. В палочках содержится пигмент родопсин, в колбочках — йодопсин. Процесс преобразования энергии в фоторецепторе начинается с поглощения фотона молекулой пигмента. Конформационное изменение молекул пигмента активирует ионы Са2+, которые посредством диффузии достигают натриевых каналов, вследствие чего проводимость для Na+ снижается. В результате снижения натриевой проводимости возникает увеличение электроотрицательности внутри фоторецепторной клетки по отношению к внеклеточному пространству. Сетчатка представляет собой довольно сложную нейронную сеть с горизонтальными и вертикальными связями между фоторецепторами и клетками. Биполярные клетки сетчатки передают сигналы от фоторецепторов в слой ганглиозных клеток и к амакриновым клеткам (вертикальная связь). Горизонтальные и амакриновые клетки участвуют в горизонтальной передаче сигналов между соседними фоторецепторами и ганглиозными клетками. #97 Охарактеризуйте особенности проводникового и коркового отделов зрительного анализатора, физиологический мех-м и значение бинокулярного зрения. Уметь определять поле зрения с помощью периметра Форстера. Зрительные пути: Аксоны ганглиозных клеток дают начало зрительному нерву. Правый и левый зрительные нервы сливаются у основания черепа, образуя перекрест, где нервные волокна, идущие от внутренних половин обеих сетчаток, пересекаются и переходят на противоположную сторону. Волокна, идущие от наружных половин каждой сетчатки объединяются вместе с перекрещенным пучком аксонов из контралатерального зрительного нерва, образуя зрительный тракт. Зрительный тракт заканчивается в первичных центрах зрительного анализатора, к которым относятся латеральные коленчатые тела, верхние бугорки четверохолмия и претектальная область ствола мозга. Латеральные коленчатые тела являются первой структурой ЦНС, где происходит переключение импульсов возбуждения на пути между сетчаткой и корой большого мозга. Нейроны сетчатки и латерального коленчатого тела производят анализ зрительных стимулов, оценивая их цветовые характеристики, пространственный контраст и среднюю освещенность в различных участках поля зрения. В латеральных коленчатых телах начинается бинокулярное взаимодействие от сетчатки правого и левого глаза. Верхние бугорки четверохолмия. Нервные клетки реагируют на движущиеся световые стимулы, включены в механизмы управления целенаправленным движением глаз. Бинокулярное зрение -механизм регуляции одновременного движения правого и левого глазных яблок, который управляются нейронами, находящимися как в подкорковых структурах, так и в коре большого мозга. Центры бинокулярного зрения находятся в области ретикулярной формации среднего мозга, в верхних бугорках четверохолмия. Ретикулярная формация среднего мозга является интегрирующим центром, получающим информацию по афферентным путям не только от верхних бугорков четверохолмия, но и от фоторецепторов сетчатки. Ядра глазодвигательных нервов находятся также под влиянием мозжечка. В мозжечке вестибулярные и зрительные сигналы интегрируются с сигналами, отражающими положение головы и глаз. Цветное зрение: восприятие глазом того или иного тона зависит от длины волны излучения: длинноволновые – красный и оранжевый; средневолновые – желтый и зеленый; коротковолновые – голубой, синий, фиолетовый. За пределами хроматической части спектра располагается невидимое невооруженным глазом ультрафиолетовое излучение. В соответствии с трехкомпонентной теорией цветового зрения нормальное ощущение цвета называется нормальной трихромазией. #98 Охарактеризуйте строение и ф-ции рецепторного, приводникового и коркового отделов слухового анализатора, методы исследования. Максимальная чувствительность слуха человека лежит в области частот от 1000 до 4000 Гц. В кортиевом органе различают внутренние и наружные волосковые клетки. Фонорецепторы кортиева органа являются вторично чувствующими рецепторами. Афферентные биполярные слуховые нейроны находятся в спиральном ганглии. От каждой клетки спирального ганглия один отросток идет на периферию к волосковым клеткам кортиева органа, а другой в составе слухового нерва направляется в ЦНС. Звук вызывает колебания эндолимфы улиткового протока попеременно в сторону вестибулярной и в сторону барабанной лестницы. Результатом такого движения является смещение основной и покровной мембраны кортиева органа относительно друг друга. Сгибание цилий является для волосковых клеток адекватным стимулом. При этом в волосковых клетках возникает рецепторный потенциал, который вызывает высвобождение медиатора. Медиатор действует возбуждающим образом на постсинаптическую мембрану афферентного волокна биполярного нейрона спирального ганглия, что в конечном счете приводит к возникновению потенциалов действия в волокнах слухового нерва. Отдельные участки улитки воспринимают определенные звуковые частоты. Каждое нервное волокно оптимально возбуждается звуком определенной частоты. У основания кортиева органа расположены рецепторные клетки, воспринимающие низкие звуки; у вершины улитки — рецепторы, воспринимающие высокие звуки. Слуховые пути: п ервичные афферентные волокна распространяются сначала к вентральной и дорсальной частям кохлеарного ядра. От вентральной части вентральный тракт направляется к ипси- и контралатеральным оливарным комплексам. Таким образом, нервные клетки в каждом оливарном комплексе получают возбуждения от рецепторов правого и левого уха, что обеспечивает сравнительную оценку акустической информации. Дорсальное кохлеарное ядро служит началом дорсального слухового тракта, волокна которого переходят на противоположную сторону и там образуют синапсы с нейронами ядра латерального лемниска. После переключения в нем слуховой тракт переключается в двух ядрах — нижнем бугорке четверохолмия и медиальном коленчатом теле. Из этих образований возбуждение распространяется к центральному концу анализатора — первичной слуховой области височной доли коры большого мозга. Подкорковые слуховые центры: первичные афферентные слуховые нейроны спирального ганглия возбуждаются чистыми тонами, т.е. очень простыми звуковыми стимулами. В противоположность этому, чем дальше от улитки по слуховому тракту находятся нейроны, тем более сложные звуковые характеристики их возбуждают. В нижних бугорках четверохолмия имеются клетки, отвечающие только на частотно модулированные тоны со специфическим направлением и различной модуляцией. Другие клетки нижних бугорков четверохолмия отвечают на тоны только в том случае, если меняется их интенсивность. Корковые центры слухового анализатора: нейронные процессы, лежащие в основе оценки звука разной частоты. Одни нейроны отвечают только на начало звукового стимула, другие — только на его окончание. Методы исследования слухового анализатора: 1. определение остроты слуха разговорной и шепотной речью; 2. исследование камертонами, воздушная проводимость, костная проводимость. 3. исследование бинаурального (пространственного) слуха.
#99 Охарактеризуйте строение и ф-ции рецепторного, проводникового и коркового отделов вестибулярного анализатора, роль статических и статокинетических рефлексов. Вестибулярный анализатор анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения тела, а также при изменении положения головы в пространстве. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры. Среди вестибулярных реакций на первом месте находятся статистические и статокинетические реакции, обеспечивающие сохранение равновесия при изменении положения тела и его частей или при возникающих ускорениях во время перемещения тела в пространстве. В осуществлении этих реакций участвуют также и проприорецепторы мышц. Рецепторы статолитовых органов и полукружных каналов: Вестибулярный орган состоит из статолитового аппарата и трех полукружных каналов, расположенных во внутреннем ухе в трех взаимно перпендикулярных плоскостях: фронтальной, сагиттальной и горизонтальной. Возбуждающим фактором для вестибулорецепторов, представленных волосковыми клетками, является наклон волосков вследствие смещения отолитовой мембраны при линейных ускорениях. Рецепторные клетки, находящиеся в ампулах, возбуждаются при угловых ускорениях вследствие движения эндолимфы по полукружным каналам. Вестибулорецепторы относятся к вторичночувствующим и связаны через синапсы с афферентными волокнами нейронов вестибулярного ганглия, расположенного в височной кости. Вестибулярные ганглии и ядра: От вестибулярных ганглиев волокна вестибулярного нерва направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, поступают к нейронам бульбарного вестибулярного комплекса: предверное верхнее ядро Бехтерева, предверное латеральное ядро Дейтерса, ядро Швальбе. Из вестибулярных ядер возбуждения направляются по вестибулоспинальному тракту к мотонейронам мышц-разгибателей; непосредственно к мотонейронам шейного отдела спинного мозга; к глазодвигательным ядрам и мозжечку; к ретикулярной формации и через таламус к задней центральной извилине коры большого мозга. Функциональные связи между вышеуказанными структурами обеспечивают не только поддержание позы человека (сохранение равновесия), но и координацию двигательных актов при выполнении целенаправленной деятельности. Статические и статокинетические рефлексы: Эти рефлексы способствуют сохранению позы, в их осуществлении большое значение имеет продолговатый и средний мозг. Статические рефлексы возникают при изменении положения тела или его частей в пространстве: 1) при изменении положения головы в пространстве — это так называемые лабиринтные рефлексы. возникающие в результате раздражения рецепторов вестибулярного аппарата; 2) при изменении положения головы по отношению к туловищу — шейные рефлексы, с проприорецепторов мышц шеи и 3) при нарушении нормальной позы тела — выпрямительные рефлексы с рецепторов кожи, вестибулярного аппарата и сетчатки глаз. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед — тонус сгибателей (лабиринтный рефлекс). Выпрямительные рефлексы — это последовательные сокращения мышц шеи и туловища которые обеспечивают возвращение тела в вертикальное положение теменем кверху. У человека они проявляются, например, во время ныряния. Статокинетические рефлексы компенсируют отклонения тела при ускорении или замедлении прямолинейного движения, а также при вращениях. Например, при быстром подъеме усиливается тонус сгибателей, и человек приседает, а при быстром спуске усиливается тонус разгибателей, и человек выпрямляется — это так называемый лифтный рефлекс. При вращении тела реакции противовращения проявляются в отклонении головы, тела и глаз в сторону, противоположную движению. Движение глаз со скоростью вращения тела, но в противоположную сторону и быстрое возвращение в исходное положение — нистагм глаз — обеспечивают сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
#100. Охарактеризуйте строение и ф-ции рецепторного, проводникового и коркового отделов обонятельного и вкусового анализаторов. Общей особенностью обонятельного и вкусового анализаторов является их способность к анализу внешних химических стимулов и формированию соответствующих обонятельных и вкусовых ощущений. Хемочувствительность рецепторов связана с высокой специфичностью и избирательностью по отношению к молекулам некоторых веществ. Анализаторы обладают большей способностью к адаптации. Постоянно действующий химический стимул достаточно быстро приводит к снижению его восприятия. Наконец, любое пищевое или непищевое вещество, попадающее в ротовую полость, неизбежно несет с собой и запаховый стимул. Обонятельные рецепторы расположены главным образом в верхней носовой раковине. Они являются первичными биполярными сенсорными клетками, имеющими два отростка: аксон и дендрит, несущий реснички. Запаховое вещество, попадая в носовую полость, вступает в контакт с мембраной ресничек. Сенсорная клетка может реагировать на несколько пахучих веществ, по которым можно построить спектр ответов одиночной обонятельной клетки. Аксоны этих клеток, направляются в обонятельную луковицу и оканчиваются на первичных дендритах отдельной митральной клетки обонятельной луковицы. Импульсы от обонятельных луковиц также поступают в гиппокамп и через амигдалярный комплекс к вегетативным ядрам гипоталамуса. Вкусовые рецепторы -специализированные сенсорные клетки, наряду с опорными и базальными клетками входящие в состав вкусовых почек. Всего у человека около 2000 вкусовых почек, которые располагаются на вкусовых сосочках языка, имеющих три разные формы: грибовидные, желобоватые и листовидные. Растворенные в воде вещества, попадающие на поверхность языка, диффундируют через пору вкусовых почек, которые образуют наружные концы сенсорных клеток. Сенсорные клетки относятся к вторичночувствующим рецепторам и отвечают на химическое раздражение формированием рецепторного потенциала. Рецепторный потенциал через синапсы вызывает возбуждение в афферентных волокнах черепных нервов, которые проводят его в мозг. Проводниковая и центральная часть вкусового анализатора. Афферентные волокна, проводящие возбуждения от вкусовых рецепторов, представлены нервом — барабанной струной (ветвь лицевого нерва), которая иннервирует переднюю и боковые части языка, а также языкоглоточным нервом, иннервирующим заднюю часть языка. Афферентные вкусовые волокна объединяются в солитарный тракт, который заканчивается в соответствующем ядре продолговатого мозга. В нем волокна образуют синапсы с нейронами второго порядка, аксоны которых направляются к вентральному таламусу. Аксоны нейронов третьего порядка проходят через внутреннюю капсулу таламуса и оканчиваются в постцентральной извилине коры большого мозга. В этой области выявлены высокоспецифичные вкусовые нейроны, реагирующие на раздражение веществами, обладающими одним вкусовым качеством. Основные вкусовые ощущения человека. У человека выявлено четыре четко различимых вкусовых ощущения: сладкое, кислое, соленое и горькое. Эти ощущения связаны со специфической чувствительностью различных участков поверхности языка. Вкус горького ощущается в первую очередь основанием языка, а сладкого — преимущественно кончиком языка. Ощущение кислого и соленого возникает при воздействии вкусовых раздражителей на боковые поверхности языка.
№ 101 Сформулируйте основные положения рефлекторной теории, охарактеризуйте безусловные и условные рефлексы и их значение для приспособительной деятельности организма. Павлов предложил рассматривать два вида поведенческих рефлексов — безусловные и условные. Безусловные рефлексы— врожденные. Безусловные рефлексы возникают на основе врожденных рефлекторных дуг. При действии адекватных раздражителей на соответствующие рецепторы безусловные рефлексы проявляются относительно постоянно. К сложным безусловным рефлексам относятся пищевые, оборонительные, половые, ориентировочно-исследовательские, родительские и др. Следует особо выделить ориентировочно-исследовательскую деятельность— реакцию животных на неожиданные, как правило, новые раздражители. Сложные безусловные рефлексы проявляются в виде специфических поведенческих реакций животных при действии на них соответствующих раздражителей. Наиболее демонстративен в этом плане сложный пищевой рефлекс. Он проявляется при действии пищи на дистантные рецепторы или на рецепторы пищеварительного тракта животного в двигательной, а также секреторной и других вегетативных реакциях — изменении дыхания, деятельности сердца и др. Сложный оборонительный рефлекс наряду с двигательной реакцией животного включает также изменение ряда вегетативных функций: секреторной деятельности пищеварительных желез, деятельности сердца, дыхания, потоотделения и т.д. Условный рефлекс — приобретаются живыми существами в индивидуальной жизни. Они связаны с обучением. Это чрезвычайно изменчивая форма рефлекторной деятельности. Ответное действие животного определяется не самим стимулом, а возникает в результате неоднократного совпадения того или иного внешнего (условного) стимула с жизненно важной деятельностью (безусловными рефлексами). Тогда ранее относительно индифферентный стимул начинает опережающе вызывать реакцию, характерную для безусловного раздражителя. Иными словами, в выработанном условном рефлексе условный стимул опережающе отражает свойства сочетанного с ним безусловного раздражителя. Непременным условием образования условных рефлексов является подкрепление, когда ранее индифферентный раздражитель неоднократно сочетается с последующим безусловным рефлексом. Другой принцип, характеризующий условнорефлекторную деятельность – принцип сигнальности. Ответная реакция организма при действии не него раздражитеоя несет в себе свойства будущего безусловного воздействия. Условный раздражитель сигнализирует о последующие безусловном рефлексе. Условные рефлексы классифицируют: - по названию условных раздражителей — световые, звуковые, обонятельные, тактильные; - по названию анализатора, воспринимающего условный раздражитель,— зрительные, слуховые, кожные; - по характеру подкрепления — пищевые, оборонительные, половые; - по методу выработки — коротко- и длительноотставленные, запаздывательные, следовые и совпадающие. При короткоотставленных условных рефлексах интервал между условным раздражителем и подкреплением обычно равен 10—20 с и не превышает 30 с. В длительноотставленных условных рефлексах этот интервал составляет более 30 с. В запаздывательных условных рефлексах интервал между условным сигналом и подкреплением равен 3 мин. В следовых условных рефлексах подкрепление предоставляется животному после прекращения действия условного раздражителя. При совпадающих условных рефлексах условный сигнал и подкрепление предоставляются животному одновременно.
№ 103 Объясните нейрофизиологические мех-мы безусловных рефлексов. Проанализируйте условия и правила выработки условных рефлексов, мех-м образования временной связи. Павлов предложил рассматривать два вида поведенческих рефлексов — безусловные и условные. Безусловные рефлексы— врожденные. Безусловные рефлексы возникают на основе врожденных рефлекторных дуг. При действии адекватных раздражителей на соответствующие рецепторы безусловные рефлексы проявляются относительно постоянно. К сложным безусловным рефлексам относятся пищевые, оборонительные, половые, ориентировочно-исследовательские, родительские и др. Следует особо выделить ориентировочно-исследовательскую деятельность— реакцию животных на неожиданные, как правило, новые раздражители. Сложные безусловные рефлексы проявляются в виде специфических поведенческих реакций животных при действии на них соответствующих раздражителей. Наиболее демонстративен в этом плане сложный пищевой рефлекс. Он проявляется при действии пищи на дистантные рецепторы или на рецепторы пищеварительного тракта животного в двигательной, а также секреторной и других вегетативных реакциях — изменении дыхания, деятельности сердца и др. Сложный оборонительный рефлекс наряду с двигательной реакцией животного включает также изменение ряда вегетативных функций: секреторной деятельности пищеварительных желез, деятельности сердца, дыхания, потоотделения и т.д. Правила выработки условных рефлексов: 1. Наличие у животного потребности и соответствующей мотивации. Например, в случае выработки пищевого условного рефлекса животное должно быть голодным. При формировании оборонительного условного рефлекса животное в ответ на повреждающее безусловное воздействие должно испытывать страх. 2. Условный раздражитель должен обязательно подкрепляться безусловным, т.е. удовлетворением жизненно важной потребности. 3. Условный раздражитель должен предшествовать подкреплению. 4. Условный раздражитель должен восприниматься животным, т.е. первоначально вызывать ориентировочно-исследовательскую деятельность. Условный раздражитель должен нести в себе экологическую значимость для животного, восприниматься им. Так, у рыб в качестве условных раздражителей более адекватными являются движение воды, изменение ее состава и др. Для высших животных это могут быть световые, звуковые, обонятельные и другие сенсорные раздражители. 5. Подкрепление по своей биологической значимости и силе должно быть сильнее условного раздражителя. 6. Условный раздражитель должен неоднократно сочетаться с безусловным подкреплением. 7. При выработке соответствующего условного рефлекса у животных должны отсутствовать конкурирующие мотивации. Например, в случае выработки пищевого условного рефлекса у животного не должен быть переполнен мочевой пузырь или животное не должно подвергаться каким-либо повреждающим воздействиям. 8. Наконец, субъект, у которого вырабатываются условные рефлексы, должен быть здоров. Механизмы образования временных: Павлов связывал образование условных рефлексов главным образом с деятельностью коры большого мозга, хотя он не отрицал участия в этом процессе и ближайших подкорковых образований. Он рассматривал процесс образования условного рефлекса как взаимодействие двух дуг возбуждений: дуги условного и безусловного рефлексов. Между этими дугами, при повторных сочетаниях образуется временная связь. Павлов обозначил ее «временной», так как при отсутствии подкрепления она быстро разрушается и условный рефлекс исчезает. Временная связь между условным раздражением и подкреплением формируется в коре большого мозга между пунктами (очагами) представительства условного сигнала и безусловного подкрепления. Образованию условнорефлекторной временной связи в коре большого мозга способствуют доминантные отношения. При этом корковый «очаг» безусловного подкрепления, будучи доминантным, притягивает к себе возбуждения, ранее вызванные условным раздражителем. Именно эти свойства способствуют образованию временной связи между пунктами условного и безусловного раздражений коры больших полушарий. Вследствие этого условный раздражитель начинает вызывать условнорефлекторный ответ.
№ 104 Объясните значение и опишите виды внешнего и внутреннего торможения в коре головного мозга. Торможение в коре головного мозга, его значение и виды: В зависимости от условий возникновения коркового торможения различают две формы – безусловное (внешнее) и условное (внутреннее). При исследовании взаимоотношений возбуждения и торможения в коре мозга было установлено, что в течение нескольких секунд после воздействия тормозного раздражителя эффект положительных условных раздражителей усиливается. И наоборот, после применения положительных условных раздражителей усиливается действие тормозящих раздражении. Внешнее (безусловное) торможение наблюдается в случаях, когда на животное с ранее выработанным условным рефлексом неожиданно действует какой-то новый, довольно сильный внешний раздражитель. Внешнее торможение проявляется также в новой обстановке. В этом случае у животного возникает ориентировочно-исследовательская деятельность, которая и является причиной торможения ранее выработанного условного рефлекса. Внешнее торможение не требует обучения. Торможение происходит в результате взаимодействия двух возбуждений. Возбуждение, обусловленное ориентировочно-исследовательской реакцией, оказывается более сильным и затормаживает более слабое возбуждение. С системных позиций при этом более сильная функциональная система на уровне отдельных нейронов мозга вытормаживает более слабую. Внутреннее (условное) торможение возникает в самой дуге условного рефлекса. Требует специального обучения. Главным условием формирования внутреннего торможения является отсутствие подкрепления условного сигнала. При этом у животных сначала, так же как и в случае внешнего торможения, в ответ на условный раздражитель развивается ориентировочно-исследовательская реакция, которая в дальнейшем сменяется отрицательной эмоцией. Различают несколько видов внутреннего торможения: Угасательное торможение развивается в тех случаях, когда условный сигнал ранее выработанного условного рефлекса перестает подкрепляться. Например, пищевой условный раздражитель при его последовательных предъявлениях перестает подкрепляться пищей. В этом случае величина слюноотделения на каждый последующий условный сигнал постепенно уменьшается и в конце концов достигает нулевых значений. Дифференцировочное торможение формируется в случае, когда один из условных раздражителей подкрепляется, а другой, близкий к нему по физическим параметрам, не подкрепляется. Торможение проявляется в этом случае по отношению к неподкрепляемому воздействию и развивается в две фазы. Сначала возникает фаза генерализации, в которой животное отвечает на оба условных — подкрепляемый и неподкрепляемый — раздражителя. Затем формируется стадия концентрации, когда на ранее подкрепляемый условный раздражитель животное отвечает условнорефлекторной реакцией, а на неподкрепляемый условный раздражитель условнорефлекторная реакция не проявляется. Запаздывательное торможение формируется в случаях, когда подкрепление отставлено от условного раздражителя, например, на 2—3 мин. При этом при пищевых запаздывательных условных рефлексах торможение проявляется в течение всего времени действия условного сигнала. Условный тормоз проявляется в тех случаях, когда условный раздражитель подкрепляется, а сочетание его с другим условным раздражителем не подкрепляется. В этом случае второй условный раздражитель становится тормозом. Механизм внутреннего торможения. Павлов считал, что внутреннее торможение развивается в корковых клетках представительства условных раздражителей. Однако опыты с регистрацией вегетативных компонентов условнорефлекторной деятельности Анохина свидетельствуют о том, что при торможении слюноотделения при пищевом условном рефлексе изменения сердцебиений и дыхания продолжают регистрироваться.
№ 105 Опишите типы высшей нервной деятельности по хар-кам основных нервных процессов, их взаимосвязь с особенностями темперамента по Гиппократу. Учение о типах поведения (темпераментах) восходит еще к Гиппократу, который по преобладанию той или иной внутренней среды подразделял людей на сангвиников, холериков, меланхоликов и флегматиков. Павлов в основу классификации типов ВНД у животных положил следующие свойства основных нервных процессов возбуждения и торможения: силу, уравновешенность и подвижность. По силе нервных процессов животных подразделяют на сильных и слабых. При этом сила процесса возбуждения характеризует предел работоспособности животных, за которым наступает пессимальное торможение. Уравновешенность отражает баланс между процессами возбуждения и торможения. Подвижность отражает способность смены процесса возбуждения на торможение и наоборот. Типы высшей нервной деятельности у собак. Павлов соотнес указанные свойства нервной системы с классификацией темпераментов по Гиппократу. Сангвиник — сильный по процессам возбуждения и торможения, уравновешенный, подвижный. Холерик — сильный, неуравновешенный (возбудимый), подвижный. Флегматик — сильный, уравновешенный, инертный. Меланхолик — слабый. Системная классификация типов высшей нервной деятельности. Системный подход изменил представления о типах ВНД человека и животных. Классификация типов при этом строится с учетом индивидуальных особенностей формирования у субъектов стадий афферентного синтеза, принятия решения, способности предвидения и оценки результатов действия. Значение различных стадий системной организации поведенческих актов меняется у одних и тех же индивидов. Человек может хорошо принимать решения в одной ситуации и быть совершенно беспомощным в другой. Индивидуальные особенности организации различных функциональных систем у каждого индивида должны учитываться при разработке и применении фармакологических и нелекарственных средств. Павлов ввел представление о двух сигнальных системах действительности. Первая сигнальная система связана с физическими свойствами условных раздражений. Она присуща животным и человеку. Вторая сигнальная система связана со словесными воздействиями на человека. Павлов рассматривал слово как «сигнал сигналов». Классификация типов высшей нервной деятельности у человека. По преобладанию первой и второй сигнальной систем Павлов подразделял людей на мыслительные, художественные и смешанные типы. У мыслительных типов преобладает вторая сигнальная система, у художественных типов — первая.
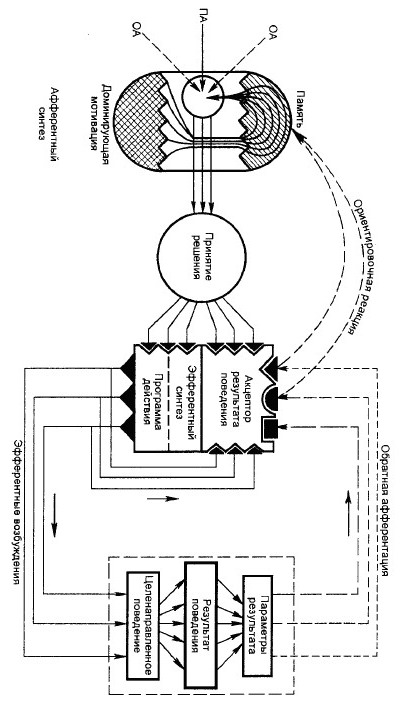
№ 106 Охарактеризуйте системную архитектонику целенаправленного поведенческого акта. ОА – обстановочная афферентация; ПА – пусковая афферентация. Центральная архитектоника поведенческого акта строится деятельностью головного мозга, являясь атрибутом сложных динамических корково-подкорковых взаимоотношений. Первой, инициативной стадией центральной архитектоники поведенческого акта является стадия афферентного синтеза, которая состоит из нескольких компонентов. Ведущим компонентом является доминирующая биологическая мотивация, которая строится на основе нервно-гуморальной сигнализации различными метаболическими потребностями. Доминирующие биологические мотивации голода, страха, жажды, полового возбуждения и др. за счет восходящих активирующих влияний специальных гипоталамических центров избирательно охватывают различные отделы головного мозга, включая кору. Биологические мотивации могут самостоятельно сформировать поведенческий акт. При этом внешние факторы играют роль ключевых, раскрывающих в определенных условиях генетические механизмы поведенческих актов. Влияния внешней среды составляют второй компонент афферентного синтеза — обстановочную афферентацию, которая непрерывно поступает в ЦНС при действии разнообразных факторов внешней среды на многочисленные экстерорецепторы живых организмов. Соотношения доминирующей мотивации и обстановки динамичны, они строятся по принципу доминанты — в первую очередь удовлетворяются биологические или обстановочные воздействия, наиболее значимые для выживания или социальной адаптации. Третьим компонентом афферентного синтеза является память. Прежде всего это генетическая память, к которой в построении поведения постоянно адресуются врожденные биологические мотивации. Механизмы памяти при определенных условиях могут самостоятельно сформировать поведенческий акт или существенно повлиять на его организацию.
№ 107 Объясните центральные мех-мы возникновения мотиваций, роль мотиваций в мех-мах афферентного синтеза. Мотивация — вызванное той или иной потребностью эмоционально окрашенное состояние организма, избирательно объединяющее нервные элементы различных уровней мозга. На основе мотиваций формируется поведение, ведущее к удовлетворению исходной потребности. Основные влечения организма представляют собой именно такую форму поведенческой деятельности, которая в отличие от рефлекторной деятельности как реакции на внешние стимулы, напротив, направлена на поиск специальных раздражителей внешней среды. К числу основных влечений, направленных на обладание определенными раздражающими предметами относятся, в первую очередь, влечение голода и ориентировочно – исследовательская деятельность. Различают биологические и социальные мотивации: Биологические мотивации, они же основные влечения, или низшие, простые, первичные мотивации. Биологические мотивации направлены на удовлетворение ведущих биологических потребностей индивидуумов по сохранению их вида или рода. К ним относятся мотивации голода, жажды, страха, агрессии, половые влечения, различные родительские, в частности материнские, температурные и другие влечения. Ведущими биологическими потребностями являются: 1) пищевая потребность, характеризующаяся уменьшением в организме уровня питательных веществ; 2) питьевая потребность, связанная с повышением осмотического давления; 3) температурная потребность — при изменении температуры тела; 4) половая потребность. Всегда имеется ведущий параметр общей метаболической потребности — доминирующая потребность, наиболее важная для выживания особи или ее рода, которая строит поведенческий акт, направленный на ее удовлетворение. Социальные мотивации, они же высшие, или вторичные, строятся на основе врожденных биологических мотиваций путем общения индивидуумов со средой обитания, родителями и окружающими их живыми существами, а у человека — и с социальной средой. В формировании социальных мотиваций значительное место принадлежит воздействию различных факторов внешней среды, обучению и, следовательно, механизмам памяти. Мотивации как биологического, так и социального плана в целостной деятельности организма всегда являются компонентом системной архитектоники поведенческого акта. Мотивации играют существенную роль в организации таких ответственных системных механизмов, как стадия афферентного синтеза, принятия решения и предвидения потребного результата — акцептора результата действия. Не меньшее значение мотивация имеет в формировании эфферентной программы поведения — стадии эфферентного синтеза. Мотивации в своем становлении в развивающемся организме подлежат общим закономерностям развития функциональных систем — процессам системогенеза.В процессе онтогенетического развития отдельных функциональных систем мотивации, будучи их составным компонентом, созревают избирательно и последовательно. Физиологические механизмы биологических мотиваций: Биологические мотивации, будучи тесно связанными с метаболическими потребностями организма, строятся на основе врожденных, генетически детерминированных механизмов. Причиной биологических мотиваций являются раздражители внутренней среды, связанные с изменением различных показателей гомеостаза, определяющих нормальное течение обменных процессов в организме, например уровня питательных веществ, осмотического давления, состояния половых клеток, различных гормонов. Биологические мотивации могут активироваться или тормозиться специальными внешними «освобождающими» факторами внешней среды, например видом или запахом противника или полового партнера, пищи или других удовлетворяющих различные потребности раздражителей. Биологические мотивации нередко относятся к разряду сложных безусловных рефлексов, или инстинктов.
№ 108 Дайте хар-ку различным видам памяти и их нейрофизиологическим мех-мам. Память — способность живых существ запоминать, сохранять и воспроизводить информацию о ранее воздействовавших на них событиях. Память тесно связана с обучением. С точки зрения системной организации физиологических функций, память присуща всем компонентам системной архитектоники поведенческого акта — стадии афферентного синтеза, принятия решения, акцептора результата действия и эфферентного синтеза. Виды памяти. Выделяют три вида памяти: кратковременную, промежуточную и долговременную. Кратковременная память определяет значимость поступающей информации для организма. Если эта информация важна для организма, особенно для удовлетворения его ведущих потребностей, она затем обрабатывается в промежуточной памяти и переходит в долговременную память. В противном случае она быстро забывается. Промежуточная память определяет сохранение полученной организмом информации в течение нескольких минут или часов. Такая память определяет, например, формирование мыслей при разговоре, запоминание адресов, телефонных разговоров, некоторых заданий (так называемая рабочая память). Долговременная память сохраняется всю жизнь. Важные для субъекта, особенно эмоционально окрашенные события, запечатляются в долговременной памяти достаточно ярко. Особую форму представляет эмоциональная память — запечатление ярких положительных и отрицательных эмоциональных переживаний. Процесс памяти включает 4 стадии: - восприятие, запечатление, запоминание информации; - хранение информации; - воспроизведение необходимой информации; - забывание. Кратковременная память формируется на основе непосредственно сенсорного отпечатка внешнего мира. При этом в памяти уже удерживается ограниченная, выделенная доминирующей мотивацией информация о внешней среде, способствующая удовлетворению ведущей потребности организма. Кратковременная память позволяет в течение нескольких секунд или минут удерживать и воспроизводить отобранную часть информации. Процессы последующей за кратковременной промежуточной памяти обычно разыгрываются в течение нескольких часов после обучения. В этот период времени экстремальные механические и химические воздействия способны стереть память, но по истечении 4 ч следы кратковременной памяти становятся устойчивыми. Происходит консолидация памяти. Кратковременная память нарушается при таких воздействиях на организм, как электрошок, сильные мозговые травмы, судороги, наркоз, гипоксия. При этом наблюдается ретроградная амнезия — потеря памяти на события, предшествовавшие воздействию. Установлено, что объем кратковременной памяти человека составляет 7±2 единицы, т.е. бессмысленные слова после однократного их применения воспроизводятся испытуемыми лишь в эквивалентном количестве. В основе современных представлений о механизмах кратковременной памяти лежит несколько гипотез: корково – подкорковая реверберация возбуждений, синаптическая теория. Долговременная память определяет сохранение ранее полученной информации в течение длительного времени. Процессы фиксации следов в долговременной памяти осуществляются лучше при повторных воздействиях, особенно биологически значимых раздражителей. Наиболее быстро процесс консолидации памяти происходит при действии эмоционально значимых раздражителей. Долговременная память по своему механизму качественно отличается от кратковременной памяти и не нарушается при таких экстремальных воздействиях на мозг, как механическая травма, электрошок, наркоз и др. Механизм долговременной памяти окончательно не установлен. Несколько теорий с разных позиций объясняют механизмы долговременной памяти: морфологические теории, глиальная теория, медиаторная теория, молекулярные теории.
№ 109 Объясните мех-мы возникновения эмоций. Проанализируйте роль положительных и отрицательных эмоций в формировании поведения. Эмоции – субъективное переживание человеком своего внутреннего состояния, в частности потребностей, а также социальных факторов окружающей среды. Системные механизмы эмоций: теория функциональных систем рассматривает эмоциональные состояния в отрезке поведенческой деятельности, включающем доминирующую потребность и ее удовлетворение, в системном кванте поведения. В этом отрезке поведенческой деятельности эмоции определяют субъективную оценку потребности, предвидение свойств потребного результата и, наконец, оценку действия на организм подкрепляющих раздражений. С позиций теории функциональных систем эмоциональная окраска поведенческого акта определяется отношением субъекта к доминирующей потребности и достигнутому результату. В случае достижения потребного приспособительного результата возникает эмоция положительного качества, биологический и социальный смысл которой — санкционировать успех поиска. В случае отсутствия достижения приспособительного результата на основе ориентировочно-исследовательской реакции нарастает общая отрицательная эмоция неудовлетворенности, которая позволяет субъекту преодолевать препятствия или строить другие формы результативной деятельности. При длительном отсутствии возможности достижения потребного результата возникает эмоциональный стресс. На основе предшествующих удовлетворений ведущих биологических и социальных потребностей эмоции все в большей степени включаются в формирование аппарата предвидения потребного результата — акцептора результата действия, определяя формирование эмоционально окрашенной цели. Эмоции, следовательно, зависят от обучения. Теории эмоций: Теория подкорковых центров: связывает генез эмоций с глубинными структурами мозга. Многочисленные клинические наблюдения указывают на то, что при локализации очага повреждения, например опухоли или кровоизлияния, в глубинных структурах мозга у пациентов наблюдаются нарушения эмоциональных переживаний от повышенной раздражительности до полного подавления эмоций. Корковая теория эмоций. Эксперименты с удалением у животных коры головного мозга отчетливо продемонстрировали, что эмоции у животных после удаления коры мозга сохраняются, но изменяют свой характер. Как правило, у таких животных эмоциональные реакции на внешние раздражители, особенно агрессивные, становятся более выраженными, яркими. Периферическая теория эмоций: в формировании эмоций существенная роль принадлежит влияниям, идущим в ЦНС со стороны внутренних органов. Интегративная (корково-подкорковая) теория эмоций: эмоции являются целостной реакцией мозга, формирующейся на основе объединения (интеграции) различных структур мозга: подкорковых образований и коры. Эндогенные эмоции. Эмоциональные возбуждения возникают первично в мотивациогенных центрах гипоталамуса, а затем генерализованно распространяются в восходящем направлении на лимбические структуры и кору большого мозга. Таким путем формируются ощущения потребностей голода, жажды, страха, полового возбуждения. Экзогенные эмоции. Эмоциональные реакции возникают под первичным влиянием внешних воздействий. В этом случае возбуждения, вызванные действием на организм внешних факторов, первично по специфическим сенсорным путям достигают клеток соответствующих проекционных зон коры большого мозга и активируют корковые механизмы памяти. Только после этого возбуждения распространяются в нисходящем направлении на эмоциогенные подкорковые и, в частности, лимбические центры, формируя в зависимости от внешних воздействий и следов памяти в одних случаях положительные, а в других — отрицательные эмоциональные реакции субъекта. Профилактика эмоционального стресса: 1. интенсивная (дозированная) мышечная нагрузка 2. микроволновое воздействие – поле УВЧ, СВЧ, определенной частоты модуляции и импульсные токи. 3. переключение на любимую работу, от которой человек получает положительные эмоции. Хобби. 4. Плавание, сауна, прием антиоксидантов, некоторых олигопептидов (вещество П, пептид, вызывающий дельта сон, бета – эндорфин, пролактин)
№ 110 Объясните физиологические мех-мы сна. Сон — физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим миром. Биологическое значение сна: активность мозга во время сна часто превосходит дневные уровни. Показано, например, что во время сна активность нейронов ряда структур мозга существенно возрастает. Во сне наблюдается и активация ряда вегетативных функций. Все это позволило рассматривать сон не как снижение процессов жизнедеятельности, а как активный физиологический процесс, активное состояние жизнедеятельности. Некоторые исследователи рассматривают сон наряду с бодрствованием как второе состояние жизнедеятельности организма. Объективные признаки сна: Потеря сознания. Сон прежде всего характеризуется потерей активного сознания субъекта, выключением его активных связей с окружающим миром. Глубоко спящий человек не реагирует на многие воздействия окружающей среды. Фазовые изменения ВНД при переходе ко сну. Особенно отчетливо фазовые состояния наблюдаются при засыпании, т.е. при переходе от бодрствования ко сну. Полиметрия. Наиболее объективно характеризуют состояние сна изменения электроэнцефалограммы и ряда вегетативных показателей. Для этого применяют методы полиметрии, когда на многоканальном полиграфе одновременно регистрируются ЭЭГ в нескольких отведениях и ряд физиологических функций: ЭКГ, дыхание, артериальное давление, температура тела, показатели газообмена. Выделяют несколько стадий изменения ЭЭГ во время сна: Медленноволновая фаза: для состояния бодрствования характерна низкоамплитудная высокочастотная ЭЭГ-активность. При закрывании испытуемым глаз и расслаблении «быстрая» бета-ЭЭГ-активность сменяется на более медленный альфа-ритм. В этот период происходит постепенное расслабление мышц, глаза закрываются, человек погружается в дремотное состояние; пробуждение в этой фазе происходит легко, достаточно слегка потревожить засыпающего. В течение следующего получаса на ЭЭГ альфа-волны начинают складываться в характерные «веретена». Стадия «веретен» примерно через 30 мин сменяется стадией высокоамплитудных медленных тета-волн. Пробуждение в эту фазу становится затруднительным. Снижаются ЧСС, кровяное давление, температура тела; сердцебиение и дыхание становятся регулярными. Стадия тета-волн сменяется стадией, когда на ЭЭГ нарастают высокоамплитудные сверхмедленные дельта-волны. Дельта – сон – период глубокого сна. Обычно при засыпании стадия медленноволнового сна занимает около 1—1,5 ч. Эта стадия сменяется появлением на ЭЭГ низкоамплитудной высокочастотной активности. Она получила по ЭЭГ-характеристике название парадоксального, или быстроволнового, сна. В состоянии быстроволнового сна испытуемые находятся в глубоком сне, их нельзя разбудить сильными раздражителями, но они просыпаются от малейшего шороха. Первое проявление парадоксального сна длится 6—10 мин. Затем на ЭЭГ снова возникают альфа-волны с последующими проявлениями фаз медленноволнового сна. Пара Дата добавления: 2015-12-15 | Просмотры: 761 | Нарушение авторских прав |