|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
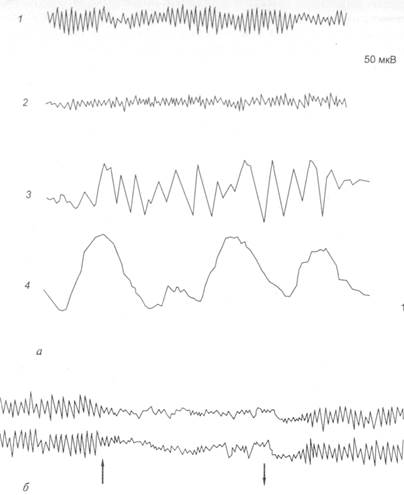
Методы исследования ВНДВажнейшим методом изучения ВНД является метод условных рефлексов в сочетании с различными дополнительными исследованиями или воздействиями. Условный рефлекс – это выработанная в онтогенезе реакция организма на раздражитель, ранее индифферентный для этой реакции. Основные правила выработки условных рефлексов следующие: неоднократное совпадение во времени действия индифферентного раздражителя с безусловным; условный стимул должен предшествовать безусловному. Следовательно, условный рефлекс образуется на базе безусловного (врожденного) рефлекса. Образованию условных рефлексов способствуют многие факторы (условия): оптимальное состояние организма (здоровье), и в первую очередь состояние коры больших полушарий; функциональное состояние нервного центра безусловного рефлекса (для пищевых рефлексов – наличие пищевой потребности); отсутствие посторонних сигналов как из внешней среды, так и от внутренних органов (кишечника, мочевого пузыря и др.); оптимальное соотношение силы условного и безусловного раздражителей. Для образования и сохранения условных рефлексов существует определенный диапазон величин подкрепления: минимальный (пороговый), оптимальный, максимальный. Широко распространенными и доступными, не травмирующими организм исследованиями деятельности мозга являются электроэнцефалография и метод вызванных потенциалов (ВП). Оба метода, казалось бы, недостаточно информативных, в последние десятилетия получили дальнейшее развитие (второе рождение) в связи с компьютерной обработкой элементов электроэнцефалограмм (ЭЭГ) и ВП. Электроэнцефалография - регистрация суммарной электрической активности мозга с поверхности головы, а электроэнцефалограмма (ЭЭГ) представляет собой кривую, зарегистрированную при этом. Запись ЭЭГ с коры головного мозга называется электрокортикограммой (ЭКоГ). Регистрация ЭЭГ производится с помощью биполярных (оба активны) или униполярных (активный и индифферентный) электродов, накладываемых симметрично в лобных, центральных, теменных, височных и затылочных областях головного мозга. Основными анализируемыми параметрами ЭЭГ являются частота и амплитуда волновой активности. На ЭЭГ регистрируется 4 основных физиологических ритма: a-, b-, q- и d- ритмы (рис. 4.1).
Рис. 4.1. Электроэнцефалограмма:
а – основные ритмы: 1 – a-ритм; 2 – b-ритм; 3 – q-ритм; 4 – d-ритм; б – реакция десинхронизации ЭЭГ затылочной области коры при открывании глаз (↑) и восстановление a-ритма при закрывании глаз (↓)
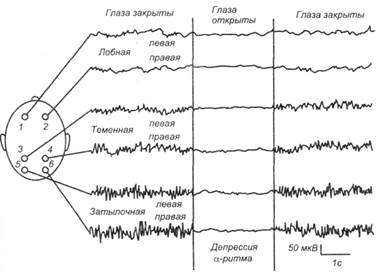
a -Ритм имеет частоту 8–13 Гц, амплитуду до 70 мкВ, наблюдается у человека в состоянии физического, интеллектуального и эмоционального покоя. a-Ритм является упорядоченным регулярным ритмом. Если он доминирует, ЭЭГ рассматривается как синхронизированная. Механизм синхронизации ЭЭГ связан с деятельностью выходных ядер таламуса (Экклс). a-Ритм преобладает у 85 –95% здоровых людей старше девятилетнего возраста. Лучше всего он выражен в затылочных областях мозга, в передних (центральной и лобной) областях, часто сочетается с b-ритмом. Вариантом a-ритма являются «веретена сна» длительностью 2 –8 с, которые наблюдаются при засыпании и представляют собой регулярные чередования нарастания и снижения амплитуды волн в частотах a-ритма. b -Ритм имеет нерегулярную частоту 14 – 30 Гц, низкую амплитуду – до 30 мкВ, сменяет a-ритм при сенсорной стимуляции, например, при действии света, при эмоциональном возбуждении. Наиболее выражен b-ритм в лобных, центральных областях головного мозга. b-Ритм отражает высокий уровень функциональной активности головного мозга. Смена a-ритма b-ритмом называется десинхронизацией ЭЭГ и объясняется активирующим влиянием на кору больших полушарий восходящей ретикулярной формации ствола и лимбической системы. q -Ритм имеет частоту 4 – 7 Гц, амплитуду до 200 мкВ. У бодрствующего человека θ-ритм регистрируется обычно в передних областях мозга при длительном эмоциональном напряжении и почти всегда регистрируется в процессе развития фаз медленноволнового сна. Отчетливо регистрируется у детей, пребывающих в состоянии неудовольствия. D -Ритм имеет частоту 0,5 – 3,0 Гц, амплитуду 200 – 300 мкВ. Эпизодически регистрируется во всех областях головного мозга. Появление этого ритма у бодрствующего человека свидетельствует о снижении функциональной активности мозга. Стабильно фиксируется во время глубокого медленноволнового сна. Происхождение q- и D-ритмов ЭЭГ связывают с активностью соответственно мостовой и бульбарной синхронизирующих систем ствола мозга. Показатели ЭЭГ. При малейшем привлечении внимания к любому стимулу развивается десинхронизация ЭЭГ - реакция блокады a -ритма (рис. 4.2). Хорошо выраженный a-ритм – показатель покоя организма (релаксации). Более сильная реакция активации выражается не только в блокаде a-ритма, но и в усилении высокочастотных составляющих ЭЭГ'. b- и g-активности. Падение уровня функционального состояния выражается в уменьшении доли высокочастотных составляющих и росте амплитуды у более медленных ритмов – q- и d- колебаний. Частотные спектры ЭЭГ помогают количественно оценивать динамику ее изменения. Выделяют несколько типов частотных спектров фоновой ЭЭГ бодрствования: ЭЭГ с a-ритмом (1) и без a-ритма (2), а также с преобладанием b-активности (3) и спектра ЭЭГ десинхронизированного типа, когда ни один из ритмов не доминирует (4).
Рис. 4.2. ЭЭГ бодрствующего человека при шести отведениях с поверхности головы
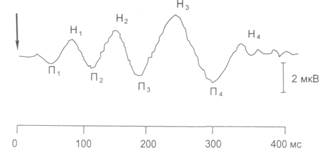
Метод вызванных потенциалов (ВП). У человека – это регистрация колебания электрической активности, возникающего на ЭЭГ при однократном раздражении периферических рецепторов (зрительных, слуховых, тактильных). У животных раздражают также афферентные пути, центры переключения афферентной импульсации, поступающей в ЦНС. Амплитуда их обычно невелика, сопоставима с волнами ЭЭГ, поэтому для эффективного выделения ВП применяют прием компьютерного суммирования и усреднения участков ЭЭГ (10 – 50), следующих до и после включения раздражающего стимула. В процессе усреднения случайные колебания ЭЭГ трансформируются в изолинию, на фоне которой отчетливо проявляются закономерные колебания ВП (рис. 4.3).
Рис. 4.3. Вызванные потенциалы у человека на вспышку света (указано стрелкой): П – позитивный ответ (гиперполяризация); Н – негативный ответ (деполяризация)
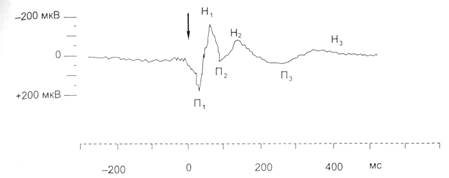
Общая продолжительность ВП составляет 300 мс. Наиболее ранние компоненты ВП отражают поступление в кору большого мозга афферентных возбуждений через специфические ядра таламуса. Эту часть ВП называют первичным ответом (рис. 4.4). Первичные ответы регистрируются в корковых проекционных зонах тех или иных периферических рецепторных зон. Поздние компоненты ВП обусловлены поступлением в кору неспецифических возбуждений через ретикулярную формацию ствола, неспецифические ядра таламуса и лимбической системы – это вторичные ответы (см. рис. 4.4). Вторичные ответы, в отличие от первичных, регистрируются не только в первичных проекционных зонах, но и в других областях мозга, связанных между собой горизонтальными и вертикальными (кора-подкорка) нервными путями.
Рис. 4.4. Первичный (П1, H1) и вторичные (П2, Н2, П3, Н3) ответы усредненного с помощью ЭВМ вызванного потенциала в соматосенсорной коре мозга кошки при электрическом раздражении лучевого нерва (указано стрелкой):
П – позитивные ответы; Н – негативные ответы
Изменения функционального состояния отражаются в ранних компонентах ВП с латентным периодом пика менее 100 мс. С привлечением внимания к стимулу, под влиянием инструкции или в результате действия электрораздражителя, который, как известно, усиливает ориентировочные реакции и сдвигает функциональное состояние в сторону возбуждения, амплитуда их растет, латентный период уменьшается. Усиление Н150 положительно коррелирует с числом ложных тревог и трудностями при выполнении задания (плохим опознанием эталонного стимула). Компонент Н150 используют в диагностических целях при определении функционального состояния с повышенной эмоциональной напряженностью. Метод дает возможность также выявить взаимодействия различных зон коры при выработке условных рефлексов. Другие методы. Магнитоэнцефалография. Поскольку при движении электрических зарядов возникает магнитное поле, мозг генерирует не только электрические (регистрируемые с помощью ЭЭГ), но и слабые магнитные волны. Напряженность этого поля более чем в раз слабее, чем у магнитного поля Земли, поэтому его можно уловить только применяя высокочувствительные датчики, заполненные жидким гелием (сверхпроводящие квантовые интерференционные устройства). Преимущество подобной методики перед ЭЭГ заключается в гораздо более высоком пространственном разрешении, т.е. повышенной точности локализации очага корковой активности, поскольку сигналы от соседних участков не накладываются друг на друга. При компьютерной томографии через мозг пропускается тонкий пучок рентгеновских лучей, источник которого вращается вокруг головы в заданной плоскости; прошедшее через череп излучение измеряется сцинтилляционным счетчиком. Таким образом получают рентгенографические изображения каждого участка мозга с различных точек. Далее с помощью компьютерных программ по этим данным рассчитывают радиационную плотность ткани в каждой точке исследуемой плоскости. В результате получают высококонтрастное изображение среза мозга в данной плоскости с пространственным разрешением 0,5 – 1 мм для слоя толщиной 2 –13 мм. Доза облучения при этом не больше, чем при обычном рентгенологическом исследовании. При позитронно-эмиссионной томографии используют радиоизотопы биологически важных атомов (18F, 15O, 13N, 11С), испускающие позитроны. Каждый позитрон, пройдя короткий (2 – 8 мм) путь, сталкивается с электроном; при этом обе частицы взаимоуничтожаются с испусканием двух g-лучей под углом 180°. Эти лучи улавливаются фотодетекторами, расположенными вокруг головы, причем их регистрация происходит лишь в том случае, когда два детектора, расположенные точно друг против друга, возбуждаются одновременно. На основании полученных данных строится изображение в соответствующей плоскости, которое отражает различия в плотности распада изотопа, т. е. в радиоактивности разных участков исследуемого объема ткани. Если изотоп включается в такие вещества, как вода, глюкоза, аминокислоты или другие биологические важные молекулы, можно изучать их распределение в мозге. Пространственное разрешение при данном методе составляет 4 – 8 мм, а временне – 1 с. Поскольку период полураспада используемых изотопов мал, поблизости от установки позитронно-эмиссионной томографии должен находиться циклотрон, на котором их получают. Ядерный магнитный резонанс. Ядра атомов вращаются, т.е. обладают моментом вращения; поскольку в их состав входят протоны, при этом генерируется магнитное поле, полюса которого расположены на оси вращения. Обычно оси вращения различных молекул распределены случайным образом, но под влиянием внешнего магнитного поля их направления меняются (подобно тому, как стрелка компаса устанавливается в магнитном поле Земли). При соответствующих условиях это приводит к резонансу атомных ядер, в результате чего они испускают электромагнитное излучение. Регистрируются появления и затухания такого резонансного излучения, которые и служат измеряемыми сигналами. Особенно хорошим резонатором является ядро атома водорода в составе воды и многих других молекул организма. В связи с этим методом ядерного магнитного резонанса в медицине измеряют именно его резонанс, причем изображения можно получать в любых плоскостях. В настоящее время толщина анализируемого слоя составляет 5–10 мм, а пространственное разрешение – около 1 мм. Однако временное разрешение пока остается слабым – 10 –20 с. Степень контрастности при ядерном магнитном резонансе такая же, как при компьютерной томографии, и, безусловно, может быть увеличена.
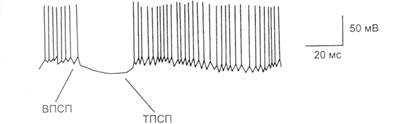
Рис. 4.5. Внутриклеточная регистрация спонтанной электрической активности моторного нейрона (клетки Беца) коры мозга с помощью стеклянного микроэлектрода. Во время ТПСП процесс генерации ПД прекращается Микроэлектродный метод основан на подведении к одиночным нейронам микроэлектродов. Чаше всего их делают в виде стеклянных микропипеток, которые перед опытом заполняются электролитом (3М КС1). Метод позволяет изучать активность одиночных нейронов ЦНС. С помощью микроэлектродов, вводимых внутрь нервных клеток, можно измерять мембранные потенциалы покоя, регистрировать постсинаптические потенциалы (возбуждающие и тормозные), а также потенциалы действия (рис. 4.5). Разновидностью микроэлектродного метода является метод микроионофореза, при котором используются многоканальные стеклянные микроэлектроды. Через один из каналов, заполненный электролитом, экспериментатор имеет возможность регистрировать электрическую активность нейрона, остальные заполняются биологически активными веществами, которые апплицируют на работающий нейрон, пропуская через растворы веществ постоянный ток. Таким образом, в условиях прямого эксперимента с регистрацией активности одиночного нейрона можно наблюдать его реакции на действие различных химических веществ и их влияние на условные рефлексы и поведение животного. Методы молекулярной биологии направлены на изучение роли молекул ДНК, РНК и других биологически активных веществ в образовании условных рефлексов. Методы холодового выключения структур головного мозга дают возможность визуализировать пространственно-временную мозаику электрических процессов мозга при образовании условного рефлекса в разных функциональных состояниях. Стереотаксический метод позволяет с помощью устройства для управляемого перемещения электродов во фронтальном, сагиттальном и вертикальном направлениях (стереотаксический прибор) ввести электрод (микропипетку, термопару и т.д.) в различные подкорковые структуры головного мозга по стереотаксическим координатам и подготовить животное для хронического эксперимента. Координаты этих структур приводятся в специальных стереотаксических атласах. Через введенные электроды можно регистрировать биоэлектрическую активность соответствующей структуры, раздражать или разрушать ее, вводить различные химические вещества. После выздоровления животного применяют метод условных рефлексов. Метод перерезки и выключения различных участков ЦНС производится механическим, электролитическим путем, а также при использовании замораживания, ультразвуковых, рентгеновских лучей. Используя электрошок или вводя снотворные вещества, можно обратимо видоизменять активность мозга в целом и наблюдать за изменением условно-рефлекторного поведения (см. также раздел 14.2). Реоэнцефалография основана на регистрации изменений сопротивления ткани мозга переменному току высокой частоты в зависимости от кровенаполнения и позволяет косвенно судить о величине общего кровенаполнения мозга, тонусе, эластичности его сосудов, состоянии венозного оттока. Эхоэнцефалография основана на свойстве ультразвука по-разному отражаться от структур мозга, его патологических образований, цереброспинальной жидкости, костей черепа и др. Кроме определения размеров локализации тех или иных образований мозга (особенно срединных) эхоэнцефалография благодаря использованию эффекта Допплера дает возможность оценивать скорость и направление движения крови в сосудах, участвующих в кровоснабжении мозга. Дата добавления: 2016-06-06 | Просмотры: 819 | Нарушение авторских прав |