|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Понятия и нейроанатомия функционального состояния организмаПонятия. Общепринятого определения функционального состояния к настоящему времени не сформировалось. Согласно нашим представлениям, функциональное состояние (ФС) – это степень активности ЦНС и других систем организма, обеспечивающих его жизнедеятельноть в различных условиях, в том числе и во время сна. Функциональное состояние оценивают обычно с двух точек зрения. Первый подход в оценке ФС – поиск и нахождение все новых и новых реакций и их комбинаций, в которых отражается динамика ФС (частота сердечных сокращений, артериальное давление, частота и глубина дыхания, скоростные характеристики двигательных реакций, изменения ЭЭЕ и др.). Однако справедливо при этом отмечается, что определение ФС через перечень различных реакций, найденных эмпирическим путем, имеет свои ограничения, так как остается невыясненным их отношение к реальным механизмам формирования функциональных состояний. Второй подход основывается на результатах трудовой деятельности (работоспособности) человека, зависящих от ФС. Следует, однако, искать нейрофизиологические критерии функционального состояния, позволяющего выполнять те или иные действия. Функциональное состояние зависит в первую очередь от активности отделов ретикулярной формации (активирующих и инактивирующих) и лимбической системы. Функциональное состояние – это психофизиологическое явление. Согласно работам В. Блока, уровень бодрствования рассматривается как поведенческое проявление различных уровней функционального состояния. Между сном и крайним возбуждением существует непрерывный ряд изменений уровня бодрствования, с которым монотонно связаны изменения в уровне активации нервных центров. Уровень активации нервных центров обусловливает уровень бодрствования.
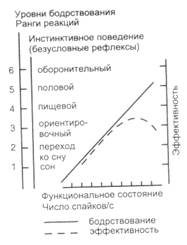
Рис. 9.1. Зависимость между функциональным состоянием, уровнем бодрствования, инстинктивным поведением (безусловными рефлексами) и эффективностью выполнения задания (по Е.Н. Соколову, Н.Н. Даниловой, 1974) Экспериментально соотношение уровня бодрствования и ФС изучалось Е.Н. Соколовым и Н.Н. Даниловой. В качестве предполагаемой меры уровня функционального состояния была взята средняя частота спайковых разрядов неспецифических и специфических нейронов таламуса кролика. Уровень бодрствования оценивался по комплексу ЭЭГ, вегетативных (дыхательных) и моторных реакций. Было найдено, что ранг уровня бодрствования линейно зависит от функционального состояния мозга, оцениваемого средней частотой спайковых разрядов неспецифических нейронов таламуса кролика (рис. 9.1). В этой схеме функциональное состояние выделено в самостоятельное явление. Показано также, что снижение уровня активации нервной системы у крыс, в частности за счет истощения в ЦНС дофамина (ДА), полностью нарушает как приобретенные при жизни навыки, так и врожденное мотивированное поведение. Если такую крысу бросить в бассейн с водой, то она погибает, так как не может плыть. Однако если ее ущипнуть за хвост или охладить воду в бассейне, что действует стимулирующим образом на животное, то крыса плывет и спасается. Это означает, что сохранность нервных связей, определяющих паттерн конкретного поведения, еще не гарантирует его реализацию (Н.Н. Данилова, А.Л. Крылова, 1999). Таким образом, механизм регуляции ФС является базальным механизмом интегративной деятельности мозга. От того, как функционирует модулирующая система мозга, зависит и обучение, и осуществление врожденного поведения. Конкретное ФС зависит от исходного уровня активности нервной системы, сохраняющей след от предшествующей деятельности субъекта. Наконец, специфика и уровень ФС существенно зависят от индивидуальных особенностей субъекта, в частности от таких его свойств, как сила–слабость нервной системы, экстроверсия – интроверсия, тревожность и т.д. Так, индивиды с сильной нервной системой менее устойчивы к монотонии: у них раньше развивается падение уровня активности нервной системы, нежели у лиц со слабым типом нервной системы. Выделяют три основные группы физиологических реакций, по которым судят об изменении ФС человека. 1. Двигательные показатели, которые измеряются количеством и интенсивностью различных фазических двигательных реакций, приходящихся на определенный интервал времени, уровень мышечного тонуса, при котором удерживается определенная поза или выполняются различные движения. 2. Вегетативные показатели -- частота (16 – 20 в минуту) и глубина дыхания, кожно-гальванический рефлекс (изменение проводимости кожных покровов), артериальное давление (120/80 мм рт. ст.), расширение и сужение сосудов головы и конечностей, частота пульса и ее дисперсия, систолический выброс (70 – 80 мл), минутный выброс сердца (4 – 5 л). 3. Показатели ЭЭГ как один из индикаторов функционального состояния организма. Нейроанатомия функциональных состояний. Ф. Бремер (1935) с помощью перерезок мозга на границе спинного и продолговатого мозга и выше моста (средний мозг) пришел к заключению, что в стволе мозга имеется структура, активирующая мозг. Во втором случае по результатам ЭЭГ наблюдалось чередование бодрствования и сна, в первом – только сон. Г. Мэгун и Дж. Моруцци (1949) показали, что электрическая стимуляция срединной части ствола мозга (ретикулярной системы) во время сна животного пробуждает его и поддерживает бодрствование. Животные с ретикулярными разрушениями становились сонными, обездвиженными и оставались такими, по крайней мере, в течение нескольких дней после операции. В ЭЭГ у них регистрировались сонные веретена, а сильные слуховые и тактильные раздражители могли вызывать лишь кратковременное пробуждение. Таким образом, приход сенсорных импульсов в кору по специфическим путям не ведет еще к длительному бодрствованию, но если ретикулярная формация сохранна, то импульсы, приходящие в нее по коллатералям от сенсорных путей, приводят к длительной активации коры. Задний гипоталамус обусловливает поведенческую активацию, а ретикулярная формация более важна для реакции пробуждения, отражающейся в изменении ЭЭГ. У человека она возникает в виде блокады a-ритма (8 – 13 в секунду) и/или усиления b-активности (14 – 30 в секунду). Повреждение заднего гипоталамуса приводит к сонливости в поведении, тогда как в ЭЭГ регистрируется высокочастотная, низкоамплитудная активность бодрствования. Таламические структуры мозга оказывают на кору двойное влияние. В режиме пачечной активности они тормозят ее, вызывая синхронизацию ЭЭГ. При одиночных спайковых разрядах кора испытывает активирующее воздействие, выражающееся в десинхронизации ЭЭГ. Полагают, что в срединной части таламуса имеется генеральный пейсмекер или ритмический осциллятор, распространяющий синхронизированные влияния на обширные области коры в виде медленных волн ЭЭГ. Реакции активации коры, вызываемые с таламуса, локальны и имеют более короткую продолжительность по сравнению с ЭЭГ-активацией, обусловленной активирующей системой ствола мозга. Они также более устойчивы к угашению, чем генерализованная ЭЭГ-активация. Таким образом, ЭЭГ-реакции активации, вызываемые активизирующей системой ствола мозга и неспецифической системой таламуса, различаются как генерализованные и локальные, тонические и фазные, быстро и медленно угасающие. Эти особенности позволяют связывать функцию активирующей системы ствола мозга с поддержанием определенного уровня активности в нервной системе, а таламическую неспецифическую систему – с селективным вниманием и локальным ориентировочным рефлексом. Нейронные пейсмекеры ритмической активности найдены в специфических ядрах таламуса (Р.А. Андерсен), в неспецифическом таламусе (Н.Н. Данилова). Фронтальная кора сдерживает чрезмерное возбуждение в ЦНС через систему ядер шва. Один из классических синдромов повреждения фронтальной коры – появление у животных двигательной гиперактивности. Предполагают, что возбуждения ретикулярной формации активируют фронтальную кору, которая, в свою очередь, через отрицательную обратную связь снижает активность ретикулярной формации. Дж. Моруцци открыл в средней части варолиева моста тормозную систему, способную вызывать сон. Под влиянием гигантоклеточного ядра моста находятся многие структуры мозга, которые и обусловливают различные тонические и фазные проявления парадоксального сна. П.К. Анохин обосновал концепцию «специфичности неспецифической активации», суть которой заключается в том, что каждый тип мотивации обеспечивается возбуждением собственной неспецифической активирующей системы, обладающей особой химической специфичностью. В опытах на животных он выделил и описал различные ЭЭГ паттерны для оборонительной, пищевой и ориентировочной мотивации и, используя различные фармакологические вещества, продемонстрировал возможность избирательной блокады каждого из этих состояний. Активирующая ретикулярная формация обеспечивает энергетическую базу исполнения реакций и имеет тенденцию участвовать в оборонительном поведении. Лимбическая система занята преимущественно активацией вегетативных процессов и связана с подготовительными фазами поведения. Функциональное состояние является результатом взаимодействия отдельных субсистем активации. Подтверждение существования двух субсистем активации, а именно, эмоциональной и неэмоциональной, получено Н.Н. Даниловой при построении семантических пространств состояний методом многомерного шкалирования (по алгоритму Торгерсона). Испытуемые ранжировали степень различия пар слов из списка, который включал названия как эмоций, так и состояний с минимальной эмоциональной компонентой (спокойствие, растерянность, уверенность и др.). Семантическое пространство состояний у всех оказалось трехмерным. Его оси ортогональны и интерпретированы как: 1) шкала знаков состояний (положительные – отрицательные состояния); 2) шкала бодрствования или готовности к действию (неэмоциональная активация – НЭА); 3) шкала выраженности эмоционального тона (эмоциональная активация – ЭА). Индивидуальные семантические пространства состояний различались субъективной значимостью трех основных осей пространства. У лиц с низкой реактивностью (по тесту Я. Стреляу) и высокой экстраверсией (по Г. Айзенку) шкала бодрствования (или неэмоциональной активации) имела более высокий удельный вес по сравнению со шкалой эмоциональной активации. ЭЭГ у животных и у человека выявила два самостоятельных волновых генератора: генератор низкочастотного ( 7– 8 Гц) и среднечастотного (9–10 Гц) a-ритма. Они были идентифицированы как две субсистемы активации - эмоциональной и неэмоциональной. Оба генератора обладают резонансными свойствами. Их активность представлена в усредненных вызванных потенциалах разными компонентами. Конкретное ФС человека в условиях бодрствования определяется взаимодействием двух систем ритмической активности. При этом чем выше активность среднечастотного a-генератора, тем ниже уровень активации низкочастотного генератора, и наоборот (Н.Н. Данилова). Оптимальность ФС для выполняемой работы связана с преобладанием активности среднечастотного α-генератора над низкочастотным. Падение эффективности выполняемой работы при высоких уровнях активации нервной системы детерминировано и преобладанием активности системы эмоциональной активации над неэмоциональной активацией. Дата добавления: 2016-06-06 | Просмотры: 629 | Нарушение авторских прав |