|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
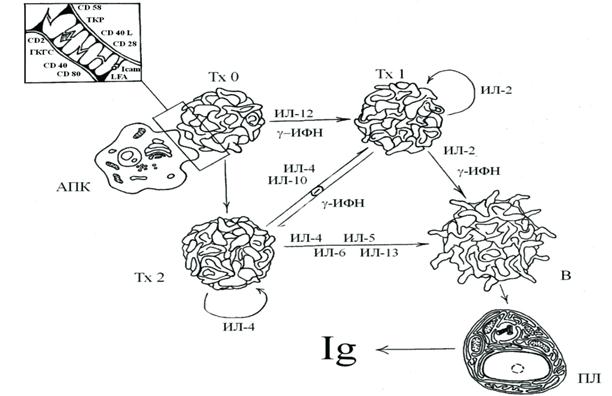
ИММУННЫЙ ОТВЕТВ настоящее время известны две формы специфического иммунного реагирования: клеточный тип иммунного ответа, осуществляемый Т-системой иммунитета, и гуморальный тип иммунного ответа, который обеспечивается В-системой иммунитета. При этом следует разделить гуморальный иммунный ответ, развивающийся на тимусзависимые антигены (или тимусзависимый гуморальный иммунный ответ) и гуморальный иммунный ответ, развивающийся на тимуснезависимые антигены (или тимуснезависимый гуморальный иммунный ответ). Клеточный иммунный ответ включает следующие основные реакции: 1. Гиперчувствительности замедленного типа (ГЗТ). 2. Реакции цитотоксических Т-лимфоцитов (или цитотоксический ответ) 3. Реакции трансплантационного иммунитета (отторжения трансплантата, хозяин против трансплантата, трансплантат против хозяина) Запуск специфического иммунного ответа осуществляется антигеном, который, чаще всего поступая в организм через барьерные ткани, контактирующие с внешней средой (слизистые оболочки, дыхательного, урогенитального, пищеварительного трактов, кожу), не уничтожается факторами неспецифической резистентности. Клетки, циркулирующие в крови, способны к активной миграции в ткани, и в зону поступления антигена, посредством хемокинов, устремляется целый поток нейтрофилов, моноцитов, здесь же накапливаются макрофаги (важную роль играет фактор, ингибирующий миграцию макрофагов). Фагоцитоз аг осуществляется преимущественно клетками-«мусорщиками» (имеют рецептор-«мусорщик»), которые не обладают специфичностью к объекту фагоцитоза. Особо стоит отметить, что макрофаги, фагоцитирующие аг в месте его проникновения, не играют важной роли в первичной обработке антигена и последующем включении Т-лимфоцитов в иммунный ответ, т.к. с одной стороны они не мигрируют в лимфоидные органы, с другой – они расщепляют аг до слишком мелких фрагментов. Антиген с током крови, лимфы или тканевой жидкости от места проникновения в организм заносится в региональный лимфоидный орган. Антигенпрезентирующие клетки (макрофаги, В-лимфоциты, дендритные клетки) захватывают, перерабатывают и представляют антигенные пептиды (т.е. осуществляют обработку аг) в комплексе с молекулами 1 или 2 класса ГКГС на своей поверхности. Макрофаги способны в основном осуществлять процессинг и презентацию корпускулярных антигенов. В-лф посредством антигенраспознающего комплекса, включающего BCR-рецептор, способны связывать и представлять растворимые молекулы. Эффективность процессинга и презентации аг АПК находится по контролем ряда цитокинов, среди основных выделяют: ИЛ-1, ФНО-альфа, ИЛ-6, ГМ-КСФ, самым мощным стимулятором макрофагов является гамма-ИФН, который усиливает экспрессию аг ГКГС 1 и 2 класса на поверхности АПК. Следующим важным этапом в развитии иммунного ответа является взаимодействие АПК с Т-лимфоцитом и распознавание антигена, что приводит в конечном итоге к активации клеток, пролиферации и дифференцировке лимфоцитов. После проникновения аг в лимфатический узел, очень быстро (в течение нескольких часов) происходит улавливание лимфоцитов, способных специфически реагировать с данным аг. Этому способствует с одной стороны усиление интенсивности кровотока через лимфатический узел и рециркуляции через него лимфоцитов, с другой – прекращение оттока из органа лимфоцитов, несущих специфические рецепторы для данного антигена. Контакт между взаимодействующими клетками осуществляется при активном участии адгезивных молекул. Решающим моментом в реализации специфического иммунного ответа является выбор пути дифференцировки CD4+-клеток в направлении Тх1 или Тх2, т.е. выбор между гуморальным и клеточным ответом. Природа антигена и определенный набор цитокинов регулируют направление дифференцировки наивных Т-х0. Этапы активации, пролиферации и дифференцировки клеток тесно взаимосвязаны. Для активации Тх0 нужно 3 сигнала: 1 сигнал: аг процессированный и презентированный на поверхности АПК в комплексе с молекулами ГКГС 2 класса. CD4+ клетки осуществляют двойное распознавание аг: TCR в комплексе с CD3 распознает сам антиген, а CD4- аг ГКГС 2 класса. Однако полноценная активация Т-х наблюдается только при взаимодействии CD28 Т-лимфоцита и CD80/86 АПК, обеспечивающим синтез ИЛ-2 (2 сигнал). Для включения третьего сигнала активации Т-х, необходима дополнительная активация АПК, которая происходит при взаимодействии CD40-рецептора АПК и CD40L CD4+-клетки. В результате активации макрофагов возрастает выработка ИЛ-1,6,10,12, ФНО, хемокинов, активных форм кислорода. При этом следует отметить, что вирусы, внутриклеточные паразиты активируют макрофаги к выработке ИЛ-12, а аллергены, внеклеточные паразиты, некоторые бактерии активируют в макрофаге синтез ИЛ-1. В В-лимфоцитах связывание молекул CD40 и CD40L вызывает переключение изотопов иммуноглобулинов в ответ на тимусзависимые антигены. Третьим сигналом для активации Тх0 служит ИЛ-1 или ИЛ-12, вырабатываемые АПК (в основном речь идет об активированных макрофагах, хотя В-лимфоциты способны синтезировать и секретировать ИЛ-1,12). Продукция цитокина ИЛ-12 индуцирует синтез гамма-интерферона Тх0 и дифференцировку их в Тх1, которые начинают секретировать цитокины (ИЛ-2,3,гамма-ИФН, ФНО-β), регулирующие развитие ГЗТ и различных цитотоксических реакций. В противоположность этому синтез ИЛ-1 индуцирует продукцию ИЛ-4, который обеспечивает дифференцировку Тх0 в Тх2. Активированные Тх2 вырабатывают цитокины (ИЛ-4,5,6,13 и др.), определяющие пролиферацию В-лимфоцитов и их дальнейшую дифференцировку. ИФН-γ негативно регулирует функцию Тх2 и, наоборот, ИЛ-4,10, секретируемые Тх2, угнетают функцию Тх1. Таким образом, результатом кооперации Тх - АПК, является взаимная активация этих клеток и выработка ими цитокинов, определяющих дальнейшее развитие иммунного ответа, а именно, пролиферацию и дифференцировку лимфоцитов.
Особая роль в этом процессе принадлежит ИЛ-2. Т-лимфоциты начинают синтезировать ИЛ-2 и одновременно экспрессировать на клеточной поверхности его рецепторы. Это обеспечивает быстрое размножение и дифференцировку антиген-специфических клонов Т-лимфоцитов.
Рис. 33. Схема гуморального иммунного ответа на тимусзависимый антиген. В настоящее время общепризнанной считается система трехкомпонентной кооперации иммуноцитов в иммуннном ответе. Это касается в первую очередь гуморального иммунного ответа на тимусзависимые антигены, который обеспечивается макрофагами, Тх и В-лимфоцитами.
Гуморальный иммунный ответ может развиваться и на тимуснезависимые антигены. Чаще всего эти антигены в своей структуре имеют повторяющиеся гомологичные эпитопы, которые образуют перекрестные сшивки с поверхностными иммуноглобулинами В-лимфоцитов и включают В-лимфоциты в процесс пролиферации и дифференцировки до зрелых антителопродуцентов.
Рис. 34. Схема формирования ЦТЛ
Реакции цитотоксических Т-лимфоцитов или цитотоксический ответ. При вирусной инфекции, опухолевом росте, отторжении трансплантата основными эффекторными клетками являются цитотоксические Т-лимфоциты (CD8+-клетки). Этапы их функционирования включают активацию, пролиферацию и дифференцировку их предшественников до зрелых эффекторов. Процесс образования ЦТЛ из наивных предшественников, покинувших тимус, осуществляется под действием двух сигналов. 1 сигнал - аг, представленный на поверхности АПК или клетках-мишенях, который пЦТЛ распознает в комплексе с антигенами ГКГС 1 класса (эффект двойного распознавания). При этом происходит экспрессия костимулирующей молекулы CD28 на ЦТЛ и активация синтеза ИЛ-2, который служит 2 сигналом активации ПЦТЛ и стимулирует их пролиферацию. В ряде случаев для дифференцировки ЦТЛ требуются дополнительные цитокиновые сигналы от Тх1 (ИЛ-2, γ-ИФН) и от других клеток (ИЛ-4,6 и др.), которые обеспечивают полноценную активацию, пролиферацию и дифференцировку пЦТЛ до зрелых ЦТЛ. Стадии иммунного ответа
Зрелые ЦТЛ осуществляют цитотоксическое действие на клетки-мишени двумя способами, вызывая либо некроз, либо апоптоз. Следующей формой клеточного иммунного ответа является реакция гиперчувствительности замедленного типа (ГЗТ). Это самая медленная форма клеточного иммунного ответа развивается на антигены, обладающие слабыми иммуногенными свойствами. Известным проявлением ГЗТ служит туберкулезная реакция – ответ сенсибилизированного организма на локальное введение туберкулина. Реакция локального воспаления развивается не только на бациллы туберкулеза, но и на широкий набор бактерий, вирусов, грибков, которые локализуются в фаголизосомах макрофагов, где они недоступны для действия как антител, так и ЦТЛ. Процессы сенсибилизации при первичном введении антигена в интактный организм осуществляются обычным путем, в результате чего формируются Тх1 (Тгзт) – сенсибилизированные лимфоциты. При повторной встрече с тем же антигеном зрелые Тх1 вступают в реакцию распознавания данного антигена, причем реакция развивается не сразу, а через 24-48 часов (отсюда ее название). Результатом распознавания является активная продукция Тх1 хемотаксических факторов, привлекающих в зону проникновения антигена макрофаги и другие клетки воспаления из кровотока. Тх1 выделяют полный коктейль цитокинов, причем для реализации ГЗТ наиболее важен гамма-ИФН, лимфотаксин, МИФ – фактор ингибирующий миграцию макрофагов, благодаря которому происходит аккумуляция макрофагов в зоне воспаления. Кооперация цитокинов вызывает сильную активацию макрофагов и стимулирует их способность убивать внутриклеточные микроорганизмы. Данный тип реакции лежит в основе многих аллергических, инфекционных, аутоиммунных заболеваний. Дата добавления: 2014-12-12 | Просмотры: 2579 | Нарушение авторских прав |