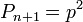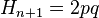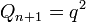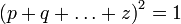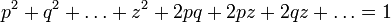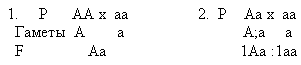|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Проблемная беседа об анализирующем скрещиванииСодержание [убрать] · 1 Комплементарность · 2 Эпистаз · 3 Полимерия · 4 См. также Комплементарность[править | править вики-текст] Основная статья: Комплементарность (генетика) Комплемента́рное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипеобусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. Прискрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлинённые. Эпистаз[править | править вики-текст] Основная статья: Эпистаз Эписта́з — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3, 7:6:3. Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7, 13:3. Полимерия[править | править вики-текст] Основная статья: Полимерия Полимери́я — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс. Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 по фенотипу происходит в соотношении 1:4:6:4:1. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1. Пример: цвет кожи у людей, который зависит от четырёх генов.
Плейотропи́я (от греч. πλείων — «больше» и греч. τρέπειν — «поворачивать, превращать») — явление множественного действия гена. Выражается в способности одного гена влиять на несколько фенотипических признаков. Таким образом, новая мутация в гене может оказать влияние на некоторые или все связанные с этим геном признаки. Этот эффект может вызвать проблемы при селективном отборе, когда при отборе по одному из признаков лидирует один из аллелей гена, а при отборе по другим признакам — другой аллель этого же гена. Содержание [убрать] · 1 Механизм · 2 Примеры o 2.1 Человек o 2.2 Другие примеры · 3 См. также Механизм[править | править вики-текст] Плейотропия — это действие одного гена на несколько фенотипических признаков. Продукт фактически каждого гена участвует как правило в нескольких, а иногда и в очень многих процессах, образующих метаболическую сеть организма. Особенно характерна плейотропия для генов, кодирующих сигнальные белки. Примеры[править | править вики-текст] Человек [править | править вики-текст] · Ген рыжих волос обусловливает более светлую окраску кожи и появление веснушек. · Фенилкетонурия (ФКУ), болезнь, вызывающая задержку умственного развития, выпадение волос и пигментацию кожи, может быть вызвана мутацией в гене, кодирующем фермент фенилаланин-4-гидроксилаза, который в норме катализирует превращение аминокислоты фенилаланина в тирозин. · Рецессивная мутация в гене, кодирующем синтез глобиновой части в гемоглобине (замена одной аминокислоты), вызывающая серповидную форму эритроцитов, изменения в сердечно-сосудистой, нервной, пищеварительной и выделительной системах. · Арахнодактилия, вызываемая доминантной мутацией, проявляется одновременно в изменениях пальцев рук и ног, вывихах хрусталика глаза и врождённых пороках сердца. · Галактоземия, вызываемая рецессивной мутацией гена, кодирующего фермент галактозо-1-фосфатуридилтрансфераза, приводит к слабоумию, циррозу печени и слепоте.
Закон Харди — Вайнберга — это закон популяционной генетики — в популяции бесконечно большого размера, в которой не действует отбор, не идет мутационный процесс, отсутствует обмен особями с другими популяциями, не происходит дрейф генов, все скрещивания случайны — частоты генотипов по какому-либо гену (в случае если в популяции есть два аллеля этого гена) будут поддерживаться постоянными из поколения в поколение и соответствовать уравнению:
Где Содержание [убрать] · 1 Статистическое обоснование закономерности · 2 Биологический смысл закона Харди — Вайнберга · 3 Условия действия закона Харди — Вайнберга · 4 Равновесие Харди — Вайнберга в реальных популяциях · 5 Практическое значение закона Харди — Вайнберга · 6 Литература · 7 См. также · 8 Ссылки Статистическое обоснование закономерности[править | править вики-текст] Рассмотрим популяцию бесконечно большого размера, в которой на частоты аллелей изучаемого гена не действуют какие-либо факторы, а также имеет местопанмиксия. Изучаемый ген имеет два аллельных состояния A и a. В момент времени (или в поколение) n, частота аллеля A = 1. 2. 3. 4. 5. 6. Потомками от скрещиваний 1, 3 и 6 будут особи с генотипами AA, Aa и aa соответственно; в результате скрещивания 2 — будет по половине особей с генотипами AA и Aa; в результате скрещивания 5 — будет по половине особей с генотипами Aa и aa; скрещивание 4 — даст все три возможных класса потомков (AA, Aa и aa) в пропорции 1: 2: 1. Исходя из вероятностей скрещиваний и пропорций в потомках от этих скрещиваний можно рассчитать частоты генотипических классов в поколении n+1.
Так как,
Аналогично можно рассчитать, что соотношение между классами P, H, Q в поколении n+2 и последующих не изменится, и будет соответствовать приведённому в начале статьи уравнению. В случае, если число рассматриваемых аллелей гена более двух, формула, описывающая равновесные частоты генотипов усложняется и её можно записать в общем виде как:
где p, q,..., z — частоты аллельных вариантов гена в исследуемой популяции. Разложив в левой части уравнения квадрат суммы получим выражение, состоящее из суммы квадратов частот аллелей и удвоенных произведений всех попарных комбинаций этих частот:
Биологический смысл закона Харди — Вайнберга[править | править вики-текст] Процесс наследования не влияет сам по себе на частоту аллелей в популяции, а возможные изменения её генетической структуры возникают вследствие других причин. Условия действия закона Харди — Вайнберга[править | править вики-текст] Закон действует в идеальных популяциях, состоящих из бесконечного числа особей, полностью панмиктических и на которых не действуют факторы отбора. Равновесие Харди — Вайнберга в реальных популяциях[править | править вики-текст] На реальные популяции в той или иной степени действуют факторы, небезразличные для поддержания равновесия Харди — Вайнберга по каким-либо генетическим маркерам. В популяциях многих видов растений или животных распространены такие явления как инбридинг и самооплодотворение — в таких случаях происходит уменьшение доли или полное исчезновение класса гетерозигот (например — см. [3]). В случае сверхдоминирования наоборот, доли классов гомозигот будут меньше расчётных. Практическое значение закона Харди — Вайнберга[править | править вики-текст] В медицинской генетике закон Харди — Вайнберга позволяет оценить популяционный риск генетически обусловленных заболеваний, поскольку каждая популяция обладает собственным аллелофондом и, соответственно, разными частотами неблагоприятных аллелей. Зная частоты рождения детей с наследственными заболеваниями, можно рассчитать структуру аллелофонда. В то же время, зная частоты неблагоприятных аллелей, можно предсказать риск рождения больного ребёнка. В селекции — позволяет выявить генетический потенциал исходного материала (природных популяций, а также сортов и пород народной селекции), поскольку разные сорта и породы характеризуются собственными аллелофондами, которые могут быть рассчитаны с помощью закона Харди — Вайнберга. Если в исходном материале выявлена высокая частота требуемого аллеля, то можно ожидать быстрого получения желаемого результата при отборе. Если же частота требуемого аллеля низка, то нужно или искать другой исходный материал, или вводить требуемый аллель из других популяций (сортов и пород). В экологии — позволяет выявить влияние самых разнообразных факторов на популяции. Дело в том, что, оставаясь фенотипически однородной, популяция может существенно изменять свою генетическую структуру под воздействием ионизирующего излучения, электромагнитных полей и других неблагоприятных факторов. По отклонениям фактических частот генотипов от расчётных величин можно установить эффект действия экологических факторов. (При этом нужно строго соблюдать принцип единственного различия. Пусть изучается влияние содержания тяжелых металлов в почве на генетическую структуру популяций определённого вида растений. Тогда должны сравниваться две популяции, обитающие в крайне сходных условиях. Единственное различие в условиях обитания должно заключаться в различном содержании определённого металла в почве). , ЗАКОН ГОМОЛОГИЧЕСКИХ РЯДОВ открытый Н. И. Вавиловым (1920) закон, согласно которому изменчивость близких по происхождению родов ивидов растений осуществляется общим (параллельным) путем. Генетически близкие роды и видыхарактеризуются сходными рядами наследственной изменчивости с такой правильностью, что зная рядформ в пределах одного вида, можно предвидеть нахождение параллельных форм у других родственныхвидов и родов. Закон гомологических рядов, как и периодическая система элементов Д. И. Менделеева вхимии, позволяет на основании знания общих закономерностей изменчивости предсказать существование вприроде не известных ранее форм с ценными для селекции признаками. Многие такие формы былинайдены после опубликования Н. И. Вавиловым закона гомологических рядов. Одним из наглядныхпримеров перспективности поиска таких форм и практического применения закона гомологических рядовявляется создание односемянных сортов сахарной свеклы. Более поздними исследованиями законгомологических рядов был подтвержден у микроорганизмов и животных, у которых обнаружен параллелизмизменчивости морфологических и биохимических признаков.
Проблемная беседа об анализирующем скрещивании. Знание законов наследования, открытых Менделем, имеет большое значение для развития сельского хозяйства и медицины. Ученые, создающие новые породы домашних животных и сорта культурных растений, подбирают родительские пары с учетом доминирования признаков, их расщепления в F2,независимого распределения. Им необходимо знать, гомо- или гетерозиготные организмы отобраны для селекционной работы. Можно ли быть уверенным, что исходные формы гомозиготны, если мы знаем лишь их фенотипы? Например, для посева вы взяли желтые семена гороха, - а какой они имеют генотип? Как узнать его? Для этой цели используют метод анализирующего скрещивания - скрещивание исследуемой особи с рецессивной исходной формой. Так, растение гороха, выросшее из желтых семян с неизвестным генотипом, скрещивают с растением, полученным из зеленых семян с известным генотипом. Так, как все половые клетки гомозиготного рецессива несут рецессивный ген (а), то характер расщепления в потомстве по фенотипу будет соответствовать качеству гамет исследуемого родителя. Если в потомстве анализирующего скрещивания отношение желтых и зеленых горошин будет 1:1, значит, генотип исследуемого родителя гетерозиготный (Аа). Если в потомстве все семена гороха имеют только желтую окраску, то исследуемый родитель гомозиготен (АА).
Из этих примеров видно, что особи, гомозиготные по доминантному гену, расщепления в F1 не дают, а гетерозиготные особи при скрещивании с гомозиготной особью дают расщеплении уже в F1. Дата добавления: 2015-02-02 | Просмотры: 954 | Нарушение авторских прав |
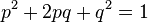
 — доля гомозигот по одному из аллелей;
— доля гомозигот по одному из аллелей;  — частота этого аллеля;
— частота этого аллеля;  — доля гомозигот по альтернативному аллелю;
— доля гомозигот по альтернативному аллелю;  — частота соответствующего аллеля;
— частота соответствующего аллеля;  — доля гетерозигот.
— доля гетерозигот.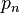 , частота аллеля a =
, частота аллеля a =  , тогда,
, тогда,  ,
,  ,
,  — частоты генотипических классов AA, Aa и aa в момент времени n. Тогда
— частоты генотипических классов AA, Aa и aa в момент времени n. Тогда 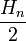 ,
,  ,
,  ,
, 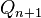 ) в следующем поколении (n+1). Возможны следующие варианты скрещивания
) в следующем поколении (n+1). Возможны следующие варианты скрещивания , вероятность
, вероятность 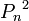
 , вероятность
, вероятность 
 , вероятность
, вероятность 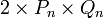
 , вероятность
, вероятность 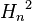
 , вероятность
, вероятность 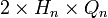
 , вероятность
, вероятность 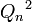
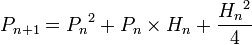
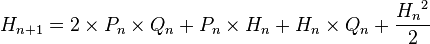
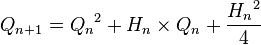
 и
и  и исходя из соотношений написанных выше между частотами аллелей а генотипических классов эти выражения можно привести к виду:
и исходя из соотношений написанных выше между частотами аллелей а генотипических классов эти выражения можно привести к виду: