|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Поступление и превращение азота в растения. Особенности усвоения молекулярного азота. Азотный обмен у растений. Усвоение других форм азота растениямиАзот составляет около 1,5% сухой массы растений. Значение азота определяется тем, что он входит в состав важных органических веществ, таких, как аминокислоты и белки, нуклеотиды и нуклеиновые кислоты, фосфолипиды, алкалоиды, многие витамины, фитогормоны (ауксины и цитокинины). Азот содержится в соединениях группы порфиринов, которые лежат в основе хлорофилла и цитохромов, многочисленных коферментов, в том числе НАД и НАДФ. Формы азота в окружающей растения среде разнообразны: в атмосфере — газообразный азот и пары аммиака, в почве — неорганические формы азота (азот аммиака, аммония, нитратов, нитритов) и органические (азот аминокислот, амидов, белка, гумуса и др.). Такое разнообразие форм азота ставило перед исследователями вопрос об источниках азотного питания для растительного организма. В растениях соединения азота также находятся в разнообразной форме. В силу этого для понимания особенностей азотного питания требовалось установить основные этапы превращения его соединений. Данная проблема имеет исключительное практическое значение. Так, наиболее часто в естественных условиях встречаются растения, содержащие недостаточное количество азота. Между тем рациональное применение азотных удобрений требует обязательного знания особенностей азотного обмена. Большая роль в выяснении всех указанных вопросов принадлежит работам академика Д.Н. Прянишникова и его учеников. Французский ученый Ж.Б. Буссенго установил, что при выращивании растений на прокаленном песке они содержат столько азота, сколько было в семени. Это доказало, что высшие растения не могут усваивать азот атмосферы. Таким образом, несмотря на то, что в атмосфере содержится около 80% азота, большинство растений не усваивают его. Однако есть растения, обогащающие почву азотом за счет атмосферы. К ним относятся представители семейства Бобовые. Г. Гельригель установил, что на корнях бобовых растений образуются вздутия — клубеньки, заполненные живыми клетками бактерий. Эти бактерии живут в симбиозе с высшими растениями и фиксируют азот атмосферы. Дальнейшие исследования показали, что фиксировать (усваивать) молекулярный азот атмосферы могут некоторые прокариотические организмы. В почве доступный для растения азот находится в основном в форме нитратов аммонийных солей. Восстановление нитратов растениями. Нитраты представляют собой окисленную форму азота и должны быть восстановлены растением до NH2, после чего они могут войти в состав аминокислот, а затем белка. Можно считать, что восстановление нитратов идет двумя путями: 1) восстановление за счет химической энергии дыхания и 2) фотохимическое восстановление в хлоропластах. Восстановление нитратов идет этапами: сначала до азотистой кислоты HNO2, затем до гидрокисламина NH2OH и, наконец, до аммиака NH3. Восстановление нитратов до NH3-и NH2-гpyпп осуществляется с помощью фермента нитратредуктазы, в состав кофермента которой входит молибден. Восстановленный азот нитратов или непосредственно поглощенный ион аммония, соединяясь с продуктами превращения углеводов, образует аминокислоты, а затем белки. Аммиак, реагируя с некоторыми органическими кислотами, может образовать аминокислоты. Так, например, аммиак, реагируя с пировиноградной кислотой, образует аминокислоту аланин:
Образовавшиеся белковые вещества подвергаются превращениям в теле растения. Животный организм все время выводит азот из своего тела в виде мочевины и отчасти мочевой кислоты. В отличие от животных растение очень бережно относится к азоту, не теряя его. При прорастании семян расщепляются запасные белки, а количество конституционных белков не только не уменьшается, а все время увеличивается. Затем происходит накопление белков в связи с переходом растения к автотрофному питанию. Роль амида, аспарагина, глютамина и мочевины в растении. При восстановлении нитратов, а также при дезаминировании аминокислот (т.е. отщеплении от них аммиака) в растениях может накопляться аммиак, который ядовит для большинства из них. В растении аммиак обезвреживается, так как он связывается аспарагиновой или глутаминовой кислотой, образуя соответственные амиды (аспарагин, глутамин). У многих низших растений образуется мочевина:
Доказан и прямой синтез мочевины из углеводов и аммиака у многих грибов (дождевики, шампиньоны). Содержание мочевины у дождевиков доходит до 10,7% от сухого вещества. Таким образом, аспарагин, глутамин и мочевина играют большую физиологическую роль, так как являются соединениями, обезвреживающими ядовитое действие аммиака, а также представляют собой резерв аминогрупп NH2 в растении для синтеза аминокислот. Подводя итоги, можно отметить два типа синтеза белков: первичный и вторичный. В обоих этих синтезах аммиак играет большую роль, что и дало возможность Д.Н. Прянишникову сказать, что аммиак есть альфа и омега (первая и последняя буквы греческого алфавита), т.е. начало и конец, превращения белков в растениях. При первичном синтезе из аммиака и углеводов строится белок (левая часть схемы). При распаде белка образуются аминокислоты, от которых при дезаминировании отщепляется аммиак, связывающийся в аспарагин или глютамин. При вторичном синтезе белков (правая и нижняя части схемы) происходит отщепление аммиака от аспарагина и образование аминокислот из углеводов (вернее, из продуктов их превращения) и аммиака. Все эти представления можно объединить в следующую схему Прянишникова:
Растительный организм, в отличие от животного, обладает способностью синтезировать все необходимые ему аминокислоты из неорганических соединений. В этом также выражается его автотрофность. Аминокислоты могут образовываться в разных органах растений — в листьях, корнях, верхушках стебля. Некоторые аминокислоты образуются непосредственно в хлоропластах и здесь используются на образование белка. Наиболее интенсивно синтез белка происходит в меристематических и молодых развивающихся тканях. Интересно, что в отрезанных листьях синтез белка полностью прекращается. Это служит еще одним доказательством, что для синтеза белка нужен какой-то фактор, образующийся в корнях растений. Можно предположить, что это фитогормон, относящийся к группе цитокининов. Для нормального протекания синтеза белка в растительном организме нужны следующие условия: 1) обеспеченность азотом; 2) обеспеченность углеводами (углеводы необходимы как материал для построения углеродного скелета аминокислот и как субстрат для дыхания); 3) высокая интенсивность и сопряженность процесса дыхания и фосфорилирования. На всех этапах преобразования азотистых веществ (восстановление нитратов, образование амидов, активация аминокислот при синтезе белка и др.) необходима энергия, заключенная в макроэргических фосфорных связях (АТФ); 4) присутствие нуклеиновых кислот. ДНК необходима как вещество, в котором зашифрована информация о последовательности аминокислот в синтезируемой молекуле белка; мРНК — как агент, обеспечивающий перенос информации от ДНК в цитоплазму; тРНК — как обеспечивающая перенос аминокислот к рибосомам; 5) рибосомы — структурные единицы, где происходит синтез белка; 6) белки-ферменты — катализаторы синтеза белка (аминоацил-т-РНК-синтетазы); 7) ряд минеральных элементов (ионы Mg2+, Са2+и др.). Образованием белка заканчивается прогрессивная ветвь азотного обмена в растениях, которая преобладает главным образом в молодых растущих органах (первичный синтез белковых веществ). Однако в растениях идет и непрерывный распад белка. Опыты с использованием меченого азота 15N позволили исследователю Ф.В. Турчину подтвердить последовательность включения азота в различные соединения, постулированную схемой Прянишникова, и одновременно показать, что обновление белка происходит чрезвычайно быстро. За 48 ч до 60% белка организма синтезируется вновь. Белки распадаются до аминокислот и далее до аммиака. Аммиак вновь обезвреживается в виде амидов (аспарагин и глутамин). На основе этих соединений образуются аминокислоты. Это позволяет организму синтезировать новый набор аминокислот, который обеспечит построение иных белков со своим специфическим набором и последовательностью аминокислот (вторичный синтез белковых веществ). Анализируя факты, Д.Н. Прянишников подчеркнул, что аммиак — это альфа и омега азотного обмена в растениях, т. е. его начальный и конечный этапы. В условиях, обеспечивающих достаточно высокий уровень синтетических процессов, аммиак представляет собой прекрасный источник азотного питания для растений. Одна из наиболее острых проблем, стоящих в настоящее время перед человечеством,- недостаток продовольствия, в первую очередь пищевого и кормового белка. Основной источник пищевого и кормового белка - растительные организмы с их уникальной способностью синтезировать белок из углекислоты, воды и неорганических соединений, но для этого растениям необходим азот. Поэтому вопросы, связанные с усвоением сельскохозяйственными культурами азота, представляют не только теоретический интерес, но и имеют огромное практическое значение. Этим вопросам во всем мире уделяется большое внимание. Периодически созываются посвященные им специальные международные конференции. На 16-й конференции Федерации европейских биохимических обществ, которая состоялась в июне 1984 г. в Москве, биохимии азотфиксации и усвоения азота растениями был посвящен специальный симпозиум. Источниками азота для растений являются: - молекулярный азот воздуха, который могут усваивать лишь некоторые почвенные микроорганизмы; - азот содержащихся в почве неорганических соединений - аммонийных солей (NH4+) и солей азотной кислоты (нитратов, NO~); - азот мочевины и различных органических соединений, образующихся в процессе разложения остатков растений и животных и их выделений (навоза, мочи). Усвоение всех этих форм азота растениями происходит благодаря действию различных ферментов. Хотя нас окружает океан молекулярного азота воздуха (N2), составляющий более 80% земной атмосферы, - однако усваивать, ассимилировать этот азот могут лишь некоторые микроорганизмы. Животные и большинство растений не способны к усвоению азота воздуха. Долгое время оставался нерешенным вопрос о возможности усвоения корневой системой растений органических форм азота. Вопрос этот можно было решить только в стерильных культурах, так как в нестерильных условиях развились бы бактерии, которые своими ферментами разложили бы органический азот и превратили бы его в минеральные формы. Корневая система высших растений находилась в простерилизованном питательном растворе, содержащем органический азот. Семена растений стерилизовались бромной водой или раствором сулемы. Опыты показали, что хотя аминокислоты и могут быть усвоены зеленым растением, но это усвоение идет крайне медленно, и растения, выращенные на этих соединениях, всегда отстают в росте от растений, получивших минеральные формы азота. Насекомоядные растения. Большой интерес представляют высшие растения со своеобразным типом азотистого питания. Сюда относятся некоторые сапрофиты, паразиты, полупаразиты и, наконец, насекомоядные растения. Своеобразие азотистого, а у некоторых форм и углеродного питания возникло в процессе эволюции под влиянием условий существования и естественного отбора. Таким образом, в отличие от грибов и бактерий, где гетеротрофное питание азотом имеет первичный характер, у этих растений оно возникло вторично. Наиболее интересную группу растений, питающихся органическим азотом, составляют насекомоядные растения. К насекомоядным растениям принадлежит примерно 500 видов растений. Все они обитатели болот. Несмотря на богатство болотных почв органическим веществом, находящийся в этих почвах органический азот недоступен для растений. Болотные почвы также очень бедны и минеральными солями (фосфор, калий и др.).
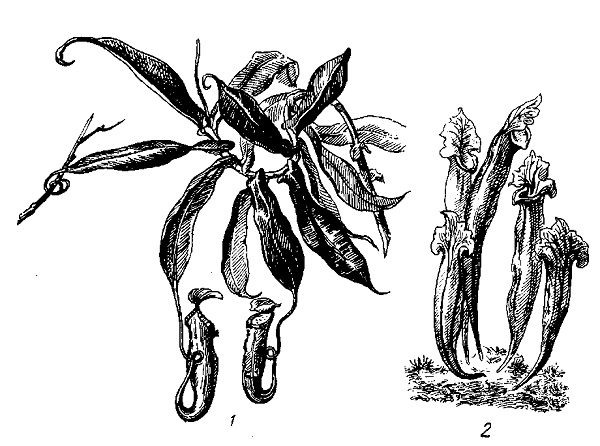
Росянка - многолетнее растение, растет на сфагновых болотах. Каждый год на поверхности мха образуется новая розетка листьев росянки. Листья снабжены многочисленными железистыми волосками (их часто называют щупальцами), выделяющими липкую жидкость, к которой и прилипают мелкие насекомые - комары и мелкие мухи. При прилипании насекомого пластинка листа свертывается. Особенно хорошо это свертывание пластинки видно у вида росянки длиннолистной. После того как насекомое прилипнет к листу, в растении начинается выделение гидролитических ферментов - протеаз, разлагающих белки, и муравьиной кислоты. Кислота способствует работе фермента протеазы, и, кроме того, она действует как яд на бактериальную флору. Последнее очень важно, так как пышное развитие флоры гнилостных бактерий могло бы сказаться отрицательно на самом растении. Ботаник Фрэнсис Дарвин, сын Чарльза Дарвина, выяснил благоприятное влияние питания росянки насекомыми. Он взял шесть сосудов с растениями росянки и разделил каждый из них перегородкой. По одну сторону перегородки росянки получали мясо, а по другую сторону им его не давали.
1 - австралийский цефалотус (лат. Cephalotus follicularis); 2 - сарацения (лат. Sarracenia) В конце опыта выяснилось, что на 100 цветков у контрольной группы, которые не получали мяса, приходится 165 цветков у получавших мясо. Иными словами, репродуктивная способность растений росянки, питавшихся мясом, сильно возрастала. Большой интерес представляет обитающая в болотистых водоемах пузырчатка. Помимо рассеченных листьев, она несет еще характерные пузыревидно измененные листья. В такой пузырек проникают мелкие рачки и уже не могут выбраться наружу, так как створка, сквозь которую проник рачок, открывается в одну сторону. Из других насекомоядных растений можно отметить кувшиноносы ( непентес) из тропиков Мадагаскара и Явы: австралийский цефалотус и американскую сарацению, листья которых имеют вид кувшинов, куда и попадают насекомые. Эти растения также выделяют гидролизирующие белки-ферменты и переваривают насекомых. У цефалотуса выделения ферментов не происходит. Многие насекомоядные растения привлекают к себе насекомых яркой окраской листьев, а некоторые выделяют сладкий сок. Так, например, у кувшиноноса в верхней части кувшинчиков имеются железки, выделяющие сладкий сок. Особенно интересна венерина мухоловка, растущая на болотах штата Каролина в Северной Америке. Это небольшое растение активно захлопывает створки листьев, когда насекомое заденет чувствительный волосок его листа.
Симбиоз и паразитизм. Особую группу покрытосеменных растений составляют сапрофиты. Встречаются они на богатой органическими веществами почве, в лесах, среди разлагающейся лесной подстилки. К ним относятся такие растения, как подъельник и орхидея гнездовка. Оба эти растения бесцветны. Правда, в листьях гнездовки содержится небольшое количество хлорофилла а, хлорофилла б у нее совсем не найдено. Подъельник - растение-сапрофит, лишенное хлорофилла. По-видимому, гриб снабжает подъельник углеводами и азотистыми веществами из малодоступного для растения перегноя, очевидно, получая от растения физиологически активные вещества (витамины), а также, возможно, и аминокислоты. Выращивая сосну в стерильных условиях и затем заражая ее определенным видом гриба (эктотрофная микориза), удалось экспериментально доказать наличие связи между высшим растением и грибом. При наблюдениях за растениями и грибами в природной обстановке выявилась тесная связь между определенными грибами и высшими растениями. По меткому выражению одного ученого, гриб кортинариус следует за березой, как "дельфин за кораблем". Большинство наших съедобных грибов образуют эктотрофную микоризу и тесно связано с определенными деревьями. Это давно отмечено в названиях грибов. В настоящее время показано, что семена орхидеи содержат очень незначительные количества витамина РР (никотиновой кислоты). Грибок снабжает семена орхидеи никотиновой кислотой, после чего они и начинают прорастать. Кроме того, синтез витамина Bi также несколько затруднен у орхидей, и снабжение семян этими веществами способствует их прорастанию и росту корней и надземной массы. Большинство травянистых дикорастущих и культурных растений также содержат эндотрофную микоризу, вызываемую низшими грибами, имеющими неразделенный перегородками мицелий. Отмечено, что при неблагоприятных условиях, например при сильном увлажнении, гриб часто становится паразитом растения. По-видимому, и в случае эндотрофной микоризы гриб снабжает растение азотом, добывая его из перегноя, а от растения получает углеводы, а также физиологически активные вещества. К последней группе растений, отклоняющихся в своем азотистом питании, относятся полупаразиты и паразиты. По-видимому, путь к паразитизму у высших растений лежал и лежит через полупаразитизм.
Много полупаразитов встречается в семействе норичниковых. Среди полупаразитов из норичниковых можно отметить характерные растения лугов: погремок, очанку и др. Растения эти присасываются своими корнями к корням других растений. Одни из них сохраняют более или менее нормальную зеленую окраску, а другие уже значительно меньше содержат хлорофилла (как, например, погремок). Основной причиной перехода этих растений к паразитическому образу жизни является слабое развитие корневой системы, вследствие чего они не могут свести своего водного баланса. Из полных паразитов можно упомянуть о видах заразихи, поражающей подсолнечник, тыквенные и ряд дикорастущих растений. Мелкие семена заразихи прорастают, стимулируемые подкислением субстрата корневыми выделениями. Основным мероприятием по борьбе с заразихой является создание невосприимчивых (иммунных) сортов. Паразит повилика в отличие от заразихи, которая поражает корни, обвивает растение и присасывается к его стеблю. У повилики имеется очень незначительное количество хлорофилла. Проросток повилики совершает круговое движение, свойственное всем растениям, но у повилики оно проявляется особенно резко. Если при этом повилика не встретит растения, вокруг которого она может обвиться, то она погибает.
Дата добавления: 2015-02-02 | Просмотры: 2055 | Нарушение авторских прав |