|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ОПЛОДОТВОРЕНИЕОплодотворение — слияние мужской (спермия) и женской (яйцеклетки) половых клеток, имеющих гаплоидный набор хромосом, в результате чего образуется новая клетка — зигота с диплоидным набором хромосом, дающая начало развитию нового организма, обладающего двойной наследственностью (матери и отца). При коитусе (половом акте) эякулят вводится в краниальную часть влагалища (крупный и мелкий рогатый скот, кролики, приматы) или проталкивается через канал шейки непосредственно в матку (свиньи, лошади, собаки). В дальнейшем продвижение спер-миев к месту оплодотворения осуществляется как за счет собственной их подвижности, так и благодаря мышечным антиперистальтическим сокращениям стенок полового тракта самки, создающим в нем отрицательное давление и эффект засасывания. Стимуляция сократительных механизмов гладкой мускулатуры матки и яйцепроводов осуществляется за счет простагландинов группы Е, присутствующих в сперме самцов, и окситоцина, выбрасываемого задней долей гипофиза самки во время полового акта. В период продвижения спермиев по половому тракту самки они подвергаются различным физическим и биохимическим воздействиям со стороны секрета матки и яйцепроводов, в результате чего в них совершаются значительные физиологические, морфологические и молекулярно-биологические изменения, приводящие к окончательному созреванию спермиев и приобретению ими оплодотворяющей способности. В процессе этих изменений, называемых реакцией капацитации, происходят удаление с поверхности спермиев слоя стабилизирующих гликопротеи-нов, приобретенного ими в придатке семенника и при взаимодействии с секретами придаточных желез во время эякуляции, изменение макромолекулярной структуры мембраны спермия, вследствие чего увеличиваются их подвижность и способность выделять
24 — 3389 из акросомной области головки протеолитические ферменты, необходимые для растворения лучистого венца яйцеклетки. Время, необходимое для завершения реакции капацитации, зависит от вида животных: у барана 1...1,5ч, у хряка 2...3, у быка и кролика 5...6, у приматов 4...5 ч после эякуляции. Продолжительность жизни и сохранение оплодотворяющей способности спермиев, находящихся в яйцепроводах крупного и мелкого рогатого скота, свиней, составляет 24...48ч, а у лошади—до 6 сут. Тем не менее старение мужских половых клеток при длительном пребывании в половом тракте самки (более 24 ч) может отрицательно сказаться на развитии и выживании зародыша, на общей плодовитости животных, поэтому очень важное значение имеет правильный выбор сроков для искусственного осеменения. Процесс оплодотворения включает ряд этапов. На первом подготовительном этапе спермин, окружившие яйцеклетку, с помощью выделяемых ими ферментов (гиалуронидазы, муциназы, трипсиноподобных ферментов и др.) деполимеризуют вещество, склеивающее клетки лучистого венца, разрыхляют его и вызывают частичное рассеивание фолликулярных клеток. После этого спермин входят в контакт с прозрачной оболочкой яйцеклетки, с помощью гликопротеидов (рецепторов) которой происходит узнавание спермиев данного вида. За редким исключением зародыши, возникающие в результате межвидового оплодотворения, не могут развиваться нормально и погибают. На следующем этапе спермин внедряются в прозрачную оболочку яйцеклетки и ее перивителлиновое простанство. У прикрепленных к прозрачной оболочке спермиев развивается так называемая акросомная реакция. В ходе ее передние две трети плазматической мембраны головки спермия сливаются с передней частью мембраны акросомы. Протеолитические и другие ферменты акросомы вступают в контакт с прозрачной оболочкой яйцеклетки, локально разрушают ее, создавая условия для продвижения спермия в глубь яйцеклетки. Только один из множества спермиев, продвигающихся к яйцеклетке и достигающих перивителлинового пространства, вступает в контакт с плазматической мембраной яйцеклетки. На этом этапе спермий сливается с плазматической мембраной и проникает в ооплазму. Хвост спермия разрушается, а головка, набухая и увеличиваясь в размерах, перемещается к центру яйцеклетки. Оболочка ядра разрушается, его хроматин разрыхляется, и после взаимодействия с белками ооплазмы ядро спермия превращается в мужской пронуклеус. Соприкосновение спермия с желточной оболочкой яйцеклетки и слияние плазматических мембран обеих гамет ведут к очень быстрым специфическим преобразованиям в яйцеклетке. За несколько секунд в ходе так называемой кортикальной реакции изменяются молекулярно-биологические свойства плазматичес- кой мембраны, а за несколько минут в ходе реакции, называемой зонной, изменяются свойства и прозрачной оболочки. Плазматическая мембрана и наружная прозрачная оболочка яйцеклетки становятся невосприимчивы к другим мужским половым клеткам, что предупреждает полиспермию — слияние яйцеклетки с несколькими спермиями. В процессе слияния яйцеклетки со спермием в ней резко активизируется обмен веществ и снимается блокада мейоза. Это ведет к завершению второго деления созревания мейоза. Хромосомы женского ядра деконденсируются и преобразуются в женский пронуклеус. Вскоре происходит первое деление дробления образовавшейся зиготы, дающее начало развитию нового организма. БЕРЕМЕННОСТЬ Беременность — особое физиологическое состояние организма самки, связанное с оплодотворением и развитием плода в матке. Беременность у коровы называют стельностью, у кобылы, ослицы, верблюдицы — жеребостью, овцы и козы — суягностью, свиньи — супоросностью, крольчихи — сукрольностью, собаки — щенностью. В зависимости от количества вынашиваемых плодов различают одноплодную и многоплодную беременность. При одноплодной беременности в матке развивается один плод, при многоплодной — несколько. У крупных животных (коров, кобыл, лосих, оленей, верблюдиц) беременность, как правило, одноплод-ная, а у мелких (свиней, овец, коз, собак, кошек, крольчих) — многоплодная. У коров двойни встречаются в 1...5 %, а тройни — в 0,1 % случаев. У кобыл двойни составляют 1...1,5 %, причем более половины из них не донашиваются. У овец рождение двоен в значительной степени зависит от породы: у тонкорунных составляет 50...80 %, у каракульских и курдючных — 10...15 (до 40 %), у овец романовской породы — до 9 %. У свиней обычно рождается 10... 12 поросят за опорос, а иногда 17...20. У собак крупных пород (овчарки, доги) рождается чаще 7... 10 (до 12) щенят, средней величины—б..^, а у карликовых пород— 2...4 щенка. У кошек рождается 2...5, у лисиц — 4, у песцов — 11, у норок — 5, у соболей — 3 детеныша. Физиологическая беременность характеризуется нормальным состоянием организма матери и рождением зрелого жизнеспособного приплода. Иногда у животных бывает добавочная беременность (суперфетация), когда беременные самки приходят в охоту и дополнительно оплодотворяются с развитием одного или нескольких новых плодов. Добавочная беременность возможна только в начальный период первичной беременности. Она считается нормальным явлением у некоторых плотоядных животных, осо-
24*
бенно у норок, которые, будучи оплодотворенными, вторично приходят в охоту несколько раз в течение полового сезона с интервалом 15...20 сут. В редких случаях подобное явление встречается у кобыл, коров, овец, свиней. При рождении нормального зрелого плода изгоняется и второй, недоношенный плод. Ложная беременность может развиваться у самок (свиньи, козы, крольчихи, собаки, кошки) после полового цикла, не сопровождавшегося осеменением, или после неплодотворного полового акта. У животных появляются признаки истинной беременности: увеличение молочных желез, молокообразование, самки подпускают чужой приплод, готовят гнездо для родов. У коз и кошек отмечены случаи скопления в матке большого количества слизи, что увеличивает контуры живота, делая их характерными для беременных животных. Через 2...3 нед признаки ложной беременности исчезают; скопившаяся в матке слизь выделяется наружу или рассасывается. Продолжительность беременности у коров составляет 270... 300 сут, у овец и коз — 145...157, у кобыл — 320...355, у свиней — ПО...118, у собак —58...66, у кошек —56...60, у крольчих —29...33, у лисиц, песцов — 49...55, норок — 37...80, нутрий — 128...137 сут. Образовавшаяся в результате слияния яйцеклетки и спермия зигота через несколько часов начинает свое развитие путем дробления в пределах прозрачной оболочки. Этот процесс начинается у свиньи через 14...16 ч, у овцы через 16...18, у коровы через 20...24, у лошади через 24 ч. Начальные стадии развития эмбриона происходят, как правило, в яйцепроводах, которые благодаря перистальтическим сокращениям своей мускулатуры, колебаниям ресничек мерцательного эпителия и обильной секреции слизистой оболочки обеспечивают постепенное передвижение зародыша к рогам матки (рис. 9.4). В рог матки зародыши поступают у свиньи через 46...48 ч, у овцы через 66...72, у коровы через 72...84, у лошади через 140...144 ч. В последующие 5...9 сут в зависимости от вида животного зародыш на стадии морулы находится в полости матки в свободном состоянии. В процессе дробления образуются два вида бластомеров: более мелкие и светлые располагаются по периферии, прилегая к прозрачной оболочке, образуют трофо-бласт (питающий листок), а более крупные и темные группируются в центре и формируют эмбриобласт (зародышевый узелок). В возрасте 5...6 сут у свиней и овец, 7 сут у крупного рогатого скота между трофобластом и эмбриобластом образуется полость — бластоцель. Таким образом, морула превращается в полый шар — бластоцисту. Клетки эмбриобласта скапливаются под трофобластом на одной стороне этого шара. Бластоциста выходит у свиней к 6...7-м суткам, у овец к 7...8-м, у лошадей к 8-м и у коров к 9...11-м суткам, как бы вылупливается из прозрачной оболочки. Происходит это вследствие истончения и разрыва ее стенки в результате роста бластоцисты, увеличения ее объема, а также выработки ею Рис. 9.4. Ранние стадии эмбрионального развития в половых путях самок млекопитающих протеолитических ферментов, вызывающих лизис стенки прозрачной оболочки. Роль наружной оболочки зародыша после этого принимает на себя трофобласт, через который осуществляется процесс интенсивного питания зародыша. Трофобласт выделяет протеолитические, гликолитические и другие ферменты, которые, оказывая воздействие на слизистую оболочку матки, обеспечивают необходимые условия для оседания зародыша и его прикрепления (имплантации) к стенке матки. Происходит это у крольчих на 7...9-е сутки после оплодотворения, у свиноматок на 11...13-е, уовцематокна 13...15-е, укоров на 15...17-е и у лошадей на 35...37-е сутки. Особенно сильно выраженными протеолитическими свойствами обладает трофобласт у приматов: при соприкосновении со слизистой оболочкой матки он вызывает растворение покровного эпителия, маточных желез, клеток стромы и сосудов слизистой и в течение 40 ч зародыш полностью погружается в слизистую оболочку матки. У сельскохозяйственных животных протеолитические свойства трофо-бласта менее выражены, и поэтому на месте соприкосновения бластоцисты со слизистой оболочкой матки отмечаются только активная гиперемия, секреция маточных желез и набухание слизистой оболочки. Однако даже при этих условиях резко улучшается питание зародыша за счет ассимиляции маточного молочка (эмбриотрофа), выделяемого железистыми клетками слизистой оболочки матки. Постепенно из эмбриобласта образуются три зародышевых листка: эктодерма, энтодерма и мезодерма, из которых развиваются все органы и ткани организма. Из эктодермы в последующем развиваются кожа, рецепторы, нервная система, начальный и конечный участки желудочно-кишечного тракта; из эндодермы — эпителий пищевода, желудка, кишечника, печень, поджелудочная, щитовидная, паращитовидная и зобная же-
лезы, эпителий трахеи, бронхов, легких, эпителий мочевого пузыря, мочеиспускательного канала; из мезодермы — гладкая мускулатура, скелетные мышцы, сердечная мышца, дерма, соединительная ткань, хрящи, сосуды, почки, семенники, яичники. Одновременно формируются и окружающие зародыш плодные оболочки, поддерживающие, защищающие эмбрион и обеспечивающие его питание (рис. 9.5). С боков эмбриона формируется складка из эктодермы и мезодермы, которая, разрастаясь, окружает эмбрион и срастается над ним. Эта внутренняя оболочка получила название «амнион». Полость амниона наполняется прозрачной водянистой жидкостью, выполняющей роль биологически активной промежуточной среды между плодом и организмом матери. Она является вначале продуктом жизнедеятельности трофо-бласта, затем продуктом секреторной деятельности амниотическо-го эпителия, а также транссудации из кровеносных сосудов амниона. Амниотическая жидкость содержит белки, жиры, углеводы, ферменты, витамины, гормоны, другие биологически активные вещества, в частности катехоламины, серотонин, гистамин, ацетил холин, простагландины, а также метаболиты всех этих веществ. Околоплодные воды являются промежуточным звеном в гормональном обмене между организмом матери и плода. Амниотическая жидкость обладает бактериостатическими свойствами. Однако не исключается возможность проникновения через амнион вирусов и даже бактерий. На ранних стадиях развития плода амниотические воды участ
ки и увлажняет родовые пути. В конце внутриутробного развития количество амниотической жидкости у овец и коз около 0,5...1,2 л, у коров 3...5, у кобыл до 3...7 л. Рис. 9.5. Схематическое изображение зародыша млекопитающего с эмбриональной плацентой и плодными пузырями (ом. — аллантоис, амн. об. — амниотическая оболочка, амн. п. — амниотическая полость, желт, мешок — желточный мешок, внезар. мез. — внезародышевая мезодерма) Наружная оболочка плода (сосудистая) — хорион возникает из трофобласта и мезодермы. Мочевая оболочка (аллантоис) образуется из выпячивания первичной кишки. У всех животных при многоплодной беременности каждый плод имеет совершенно обособленные водную, мочевую и нередко сосудистую оболочки. У коров при многоплодной беременности сосудистая оболочка часто бывает общей, но с обособленными для каждого плода зонами распространения кровеносных сосудов и плацентарных связей. При образовании анастомозов между сосудами различных плодов кровь одного плода попадает в кровеносную систему другого, оказывая на него определенное эндокринное воздействие. Гормоны мужской особи задерживают нормальное развитие половых органов у телочек, и они, как правило, рождаются с недоразвитыми половыми органами и остаются бесплодными. У овец и коз при многоплодной беременности общая сосудистая оболочка нередко встречается, но анастомозы кровеносных сосудов отмечаются как исключение. Внутриутробный период развития разделяют на три основные фазы. Зародышевая фаза (до имплантации эмбриона) продолжается 7...15 сут (7...9 у крольчих, 11...13 у овцематок и свиноматок, 13...15 у коров и кобыл). Эмбриональная фаза — фаза закладки органов продолжается у крольчих с 9-х по 18-е сутки, у овцематок с 13-х по 46-е, у свиноматок с 13-х по 38-е, у коров с 13-х по 60-е и у кобыл с 15-х по 97-есутки. Плодная фаза — фаза структурного и физиологического совершенствования органов продолжается у крольчих с 19-х по 30-е сутки, у овцематок с 47-х по 150-е, у свиноматок с 39-х по 114-е, у коровы с 61-х по 284-е и у кобыл с 98-х по 335-е сутки. Эмбриональная диапауза. У некоторых видов животных зигота, достигнув стадии бластоцисты, длительное время не прикрепляется к слизистой оболочке рогов матки (куницеобразные, косуля, медведи, некоторые виды кенгуру, броненосцы, скунсы, морской заяц). Период от оплодотворения ооцита до имплантации зародыша получил у этих животных название эмбриональной диапаузы или латентной стадии беременности. Эмбриональная диапауза имеет большое биологическое значение, позволяя животным рожать потомство в наиболее благоприятный сезон года. Например, у соболей сезон спаривания приходится на июль, а щенение самок происходит в апреле — начале мая следующего года. Тем не менее в феврале и начале марта бластоцисты в матке находятся еще в свободном состоянии. Следовательно, беременность от начала имплантации бластоцист до родов длится у соболя около 1 мес. Зигота у американской норки достигает в своем развитии стадии бластоцисты за 8 сут и может находиться в роге матки в таком состоянии до 49 сут. Заканчивается эмбриональная диапауза за 29 сут до родов. Во время эмбриональной диапаузы желтые тела представляют собой вполне
сформированные образования, но уровень их секреторной деятельности ниже, чем при имплантации.
Плацента. В связи с имплантацией и развитием зародыша начинают происходить глубокие изменения в матке. Уже на ранних стадиях развития трофобласт плотно прилегает к эндометрию и внедряется в него. Вначале это очень рыхлая связь, но постепенно благодаря изменениям в эндометрии, хорионе, как и позднее в аллантохорионе контакт становится теснее (имплантация). Возникшая при этом структура называется плацентой, она предназначена для связи плода с материнским организмом и осуществления обмена веществ между ними. Это решающий фактор эмбриогенеза, определяющий нормальное развитие плода. Плацента является уникальным образованием, выполняющим одновременно функцию легких, кишечника, печени, почек и эндокринной железы. Плацента имеет материнскую часть, образовавшуюся из эндометрия, и зародышевую, образовавшуюся из хориона или алланто-хориона и обращенную в полость плода. По расположению ворсинок хориона различают четыре типа плацент (рис. 9.6). Рассеянная, или диффузная. Ворсинки равномерно покрывают всю поверхность хориона. Этот тип встречается у лошадей, свиней, ослиц, верблюдиц. Множественная, или котиледонная. Ворсинки образуют группы на отдельных участках — котиледоны, которые соединяются с утолщениями матки — карункулами. Этот тип встречается у коров и овец. Зональная (кольцевидная, или поясковая). Ворсинки опоясывают среднюю часть хориона, образуя структуру, напоминающую кольцо. Характерна для самок плотоядных животных (кошки, собаки). Дисковидная. Ворсинки сосредоточены на ограниченной поверхности хориона. Дисковидные плаценты характерны для всех представителей грызунов и приматов (в том числе для человека). По глубине проникновения ворсинок хориона в материнскую часть плаценты, т. е. по степени взаимосвязи плодной и материнской ткани, плаценты подразделяют также на четыре типа (рис. 9.7). Эпителиохориальная (полуплацента) плацента имеется у кобыл, свиней, верблюдиц, ослиц, самки бегемота и неко-
Рис. 9.7. Взаимоотношения зародышевых и материнских тканей в плацентах различных типов: А — эпителиохориальная плацента (свинья); Б — десмо-хориальная плацента (жвачные); В— эндотелиохориаль-ная плацента (хищные); Г — ворсинковая гемохориаль-ная плацента (приматы); / — трофобласт; 2—соединительная ткань хориона с зародышевыми сосудами; 3 — эпителий матки; 4 — соединительная ткань слизистой оболочки матки с материнскими сосудами; 5— кровеносные лакуны торых сумчатых. Ворсинки хориона проникают в просвет маточных желез, тесно соприкасаясь со слизистым эпителием матки, но не разрушают материнские ткани. Питание плода после имплантации осуществляется за счет маточного молочка или эмбриотро-фа. При родах не происходит разрушения и отторжения ткани эндометрия; кровотечения нет. Десмохориальная (синдесмохориальная, соединитель-нотканая) плацента имеется у жвачных (корова, овца, коза, лосиха). Ворсинки хориона разрушают эпителий и железы эндометрия, внедряясь в соединительную ткань стенки матки — карунку-лы. При родах целостность эндометрия нарушается только на отдельных участках. Эндотелиохориальная (вазохориальная) плацента имеется у хищных животных. Ворсинки хориона, внедряясь, разрушают не только эпителий и соединительную ткань эндометрия, но и стенки кровеносных сосудов, кроме эндотелия. Питание плода происходит из крови матери, отделенной от трофобласта толь- ко эндотелием материнских сосудов. Этот более совершенный тип плацентации обеспечивает снабжение плода питательными веществами и кислородом. При родах отторгается только часть эндометрия; кровотечение не особенно сильное. Гемохориальная плацента имеется у некоторых насекомоядных, летучих мышей, грызунов, приматов (обезьяны, человек). Ворсинки хориона разрушают ткани и кровеносные сосуды эндометрия и омываются непосредственно кровью матери. При этой связи устанавливается более тесный контакт плода с материнским организмом. При родах происходит отторжение эндо-метриального слоя матки; кровотечение обильное. Функции плаценты. Плацента — сложный по своим функциям орган, через который осуществляются питание и газообмен плода, выделение продуктов метаболизма, формирование его гормонального и иммунного статуса. Плацента выполняет не только транспортные функции, но и сама синтезирует необходимые для плода питательные и биологически активные вещества, включая гормоны. Плод получает от матери через плаценту кислород, питательные вещества, воду, витамины, электролиты, антитела, а передает матери углекислоту и конечные продукты обмена веществ. Обмен кислорода и растворимых веществ через плацентарную мембрану подчинен законам осмоса и диффузии, чему способствует разница парциального давления в крови матери и плода. В крови плода концентрация калия, натрия, фосфора выше, а белков, липидов, нейтральных жиров, глюкозы ниже, чем в крови матери. Газообмен в плаценте аналогичен процессу в легких и определяется наличием плацентарного кровообращения. Транспорт газов происходит постоянно, так как плацента не обладает способностью к накоплению кислорода и углекислоты. В плаценте обнаружены витамины А, В, С, D и многие ферменты, под действием которых белки, жиры, углеводы расщепляются и могут усваиваться тканями плода. В ней откладываются запасы гликогена, жира и витаминов, т. е. в плаценте не только синтезируются, но и депонируются питательные вещества. Через плаценту не проникают в кровь плода крупномолекулярные вещества, некоторые микроорганизмы, паразиты и их яйца. Способность эпителия хориона пропускать одни вещества из крови матери в кровь плода и задерживать или подвергать биохимической обработке другие получила название плацентарного барьера. Однако возможен переход из крови матери некоторых вредных веществ, таких, как наркотики, алкоголь, никотин, сульфаниламиды, антибиотики, ртуть, мышьяк и т. д. Вирусы также способны преодолевать плацентарный барьер. Во взаимоотношениях между организмом матери и плода плацента выполняет роль эндокринного органа. В плаценте всех млекопитающих животных синтезируется эстрадиол. В плаценте приматов и овцы синтезируется прогестерон, причем в таком количе- стве, что плацента полностью заменяет функцию желтого тела яичника, которая прекращается у человека на четвертом месяце беременности, у овцы — в конце второго месяца. У коз, свиней и коров плацента также вырабатывает прогестерон, но в небольшом количестве, не достаточном для сохранения беременности после экспериментального удаления желтого тела. В плаценте приматов и грызунов синтезируется хорионический гонадотропин, который в ранние сроки беременности способствует развитию и становлению функциональной активности желтого тела. В плаценте лошадей синтезируется гонадотропин, действующий на организм аналогично фолликулстимулирующему гормону гипофиза. Он вырабатывается особыми структурами, локализованными в слизистой оболочке матки, — эндометриальными чашами, образованными клетками хориона. Доказательств выработки гонадотропина плацентой у других животных пока нет. Вместе с тем у овец, коз, самок крупного рогатого скота и приматов плацента синтезирует другой тип гормона белковой природы — плацентарный лактоген-ный гормон, обладающий лютеотропным действием, т. е. стимулирующий функционирование желтого тела. Значительна роль плаценты в иммунологическом обеспечении развития беременности. Биологически активные вещества и гормоны, синтезируемые плацентой, оказывают влияние на различные отделы иммунной системы матери, подавляют активность лимфо-поэза, блокируют рецепторы воспринимающих клеток. Особенности плацентарного кровообращения и кровообращения плода. Питательные вещества плод при плацентарном кровообращении получает посредством сосудов пуповины. Обмен веществ происходит между материнской кровью, циркулирующей в сосудах слизистой оболочки матки (материнская плацента), и кровью плода, циркулирующей в капиллярах его сосудистой оболочки (плодная плацента). Кровообращение плода представляет собой замкнутую систему, изолированную от кровеносной системы матери; кровеносные сосуды матери и плода между собой не сообщаются (рис. 9.8). Все органы и ткани тела плода снабжаются кровью, содержащей небольшое количество кислорода и повышенное — углекислоты. Кровь плода, получившая питательные вещества и кислород из крови матери, приближается по показателям к артериальной. Из многочисленных капилляров сосудистой оболочки (хориона) плода формируется крупный сосуд — пупочная вена (у жвачных —двойная). Она находится в пупочном канатике и через пупочное кольцо брюшной стенки поступает в плод, затем к воротам печени и соединяется с воротной веной, где происходит первое смешение артериальной крови пупочной вены с венозной кровью воротной вены. У плодов лошади и свиньи смешанная кровь распространяется по капиллярам печени, собирается в печеночную вену, впадающую в заднюю полую вену, где смешивается с венозной кровью
1 — плацента; 2 — вена от плаценты; 3 — пупочные артерии от подвздошной артерии плода; 4— печень; 5—венозный проток; б—задняя полая вена; 7—передняя полая вена; 8— правое предсердие; 9—овальное отверстие между предсердиями; 10— легочная артерия; 11 — ветви, питающие легкие; 12— боталлов проток; 13— дуга аорты; 14— ветви от подвздошных артерий, переходящие в пупочные артерии (густота штриховки соответствует степени венозности крови) задней полой вены. У плодов жвачных животных пупочная вена минует печень. Кровь пупочной вены через особый (аран-циев) проток поступает в заднюю полую вену. Небольшая часть крови из пупочной вены проникает через воротную вену в печень для питания этого органа. По задней полой вене смешанная венозная кровь поступает в правое предсердие. У плода в перегородке между предсердиями находится овальное отверстие с клапаном, открывающимся в сторону левого предсердия. В правом предсердии смешанная кровь, поступающая по задней полой вене, еще раз смешивается с венозной кровью, поступающей по передней полой вене. Из правого предсердия часть крови проникает через овальное отверстие в левое предсердие, затем в левый желудочек, а из него в аорту. Небольшая часть крови попадает в правый желудочек, а оттуда в легочные артерии. Легкие у плода не функционируют, поэтому кровь из легочной артерии, минуя легкие, поступает непосредственно в аорту по ботал-ловому протоку и течет по большому кругу кровообращения, обеспечивая питание всего организма. После отдачи питательных веществ тканям плода венозная кровь собирается в полые вены. От разветвления тазовых артерий отходят две парные пупочные артерии, которые вместе с мочевым протоком проходят в пуповине и, доходя до сосудистой оболочки (хориона) плода, дробятся там на мелкие ветви и капилляры, из которых кровь собирается в сосуды, образующие пупочные вены. Таким образом замыкается круг плацентарного кровообращения. Функциональные изменения в организме беременных самок. Развитие беременности сопровождается рядом существенных изменений не только в половой системе самки, но и в деятельности других органов. Меняются интенсивность и характер обменных процессов. Повышается аппетит, увеличиваются потребление кормов, пищевое возбуждение, секреторная деятельность желез пищеварительного аппарата, укорачивается время переваривания корма. В обменных процессах начинают преобладать процессы ассими- ляции. Происходит накопление азота, жира, гликогена, минеральных веществ. Возрастает потребность в питательных и минеральных веществах, витаминах. В крови повышается количество жирных кислот, нейтрального жира, фосфолипидов, холестерина. У беременной самки на 15...20% повышается основной обмен. Увеличивается масса животного. В крови снижается уровень кальция и фосфора, усиливается извлечение их из костей. Усиливается кроветворение, в крови повышается число эритроцитов и лейкоцитов, количество гемоглобина. Изменяется функциональная деятельность сердечно-сосудистой системы. Усиливается работа сердца, увеличивается минутный объем крови. Происходит физиологическая гипертрофия сердечной мышцы (особенно левого желудочка); возможна тахикардия. Повышается тонус сосудов. Несколько учащается дыхание. Большую функциональную нагрузку испытывают печень и почки. Гипертрофируются мышцы брюшной стенки, развиваются молочные железы, увеличивается объем вымени. К концу беременности увеличивается подвижность в сочленениях таза. Благодаря перестройке обмена веществ и деятельности органов создаются условия для поступления веществ к плоду, для его роста и развития. Во время беременности в организме самки увеличивается объем циркулирующей крови, усиливается эритропоэз, повышается свертываемость крови, возрастает скорость оседания эритроцитов (СОЭ). Гипофиз становится больше за счет передней доли; повышаются образование и концентрация в крови гормонов передней доли гипофиза — пролактина, лютеинизирующего гормона, адренокор-тикотропного, тиреотропного, соматотропного гормона. Гипертрофируются надпочечники, в крови повышается концентрация их гормонов — кортикостероидов, катехоламинов. Увеличивается щитовидная железа, повышается ее функция (в первой половине беременности), в крови повышается концентрация ее гормонов — тироксина и трийодтиронина. Островковый аппарат поджелудочной железы начинает больше выделять инсулина. Повышается тонус парасимпатической, а позже симпатической нервной системы. К концу беременности понижается возбудимость коры больших полушарий головного мозга, но повышается возбудимость спинного мозга. Для нормального протекания беременности чрезвычайно важно поддержание определенного гормонального фона в организме самки. При половом цикле, не закончившемся беременностью, с 10... 12 сут начинается регрессия желтого тела; концентрация прогестерона в отходящих от яичников сосудах снижается. В случае же оплодотворения этого не происходит, что объясняется воздействием на гормональный фон самих зародышей. Один из установленных факторов их влияния — выработка ими значительного количества эстрогенов на 12-е сутки после осеменения, т. е. непо-
средственно перед имплантацией. Благодаря этому обеспечивается сохранение желтого тела. Для поддержания беременности концентрация прогестерона не должна падать ниже нормального уровня в течение всей беременности. РОДЫ Роды — сложная реакция организма беременной самки, возникающая в связи с завершением развития плода и выражающаяся в выведении из матки плода и плодных оболочек. Родам предшествуют определенные структурно-физиологические изменения в организме самки. Эти изменения являются предшественниками родов, начинают проявляться за 1...3 сут до родов: расслабление связочного аппарата таза (крестцовые кости приобретают возможность смещаться вверх и вниз); опухание и увеличение половых губ; отвисание живота; укорочение шейки матки; разжижение густой, липкой слизи влагалища, слизистой пробки^ закрывающей канал шейки матки, и выделение этой слизи из наружных половых органов; увеличение молочной железы и выделение из нее секрета. Родовой процесс условно делят на три последовательные стадии: подготовительная, выведения плода и выведения последа. Подготовительная стадия начинается сокращениями матки — родовыми схватками, чередующимися с расслаблениями. Схватки вначале короткие, с паузами до 30 мин, затем интервалы уменьшаются до 3...5 мин. Сокращения начинаются в связках, поддерживающих матку, затем переходят на вершину рога и перемещаются дальше к шейке матки. При сокращении матки в ней перемещаются мышечные пласты, она утолщается и подтягивается к позвоночнику. Раскрывается канал шейки матки. Изменяются^ позиции и расположение плода. Плод поворачивается на 90—180° за счет волнообразных движений плодной жидкости, вызванных сокращениями матки. Движется и сам плод. Под давлением плодные оболочки проникают в канал шейки матки, разрывается сосудистая оболочка и через разрыв выходят мочевая и водная оболочки, которые расширяют шейку матки. Плодные оболочки разрываются, и околоплодная жидкость изливается, увлажняя родовые пути и подготавливая их к изгнанию плода. Часть околоплодной жидкости задерживается плодом и выливается позже. Стадия выведения п л о д а — продвижение плода по родовому каналу начинается активными сокращениями матки, мышц брюшного пресса и спинных мышц. Сокращения мышц брюшного пресса и спинных мышц, связанные с выведением плода из матки, называются потугами. Потуги продолжительностью около 1 мин чередуются паузами до 2...3 мин. За счет движущей силы — схваток и потуг плод выводится через родовые пути. Стадия выведения последа наступает после рождения плода. В эту стадию схватки и потуги становятся реже. Понижение давления в ворсинках сосудистой оболочки, связанное с родами, ослабляет связь околоплодных оболочек с материнской частью плаценты. За счет схваток и потуг плодная часть плаценты отделяется от материнской части и выводится через родовые пути. У коров в связи с особенностями связи плодной и материнской части плаценты стадия выведения последа более продолжительная, чем у других видов животных; отмечаются случаи задержания последа, требующие врачебного вмешательства (табл. 9.4). Дата добавления: 2015-12-15 | Просмотры: 936 | Нарушение авторских прав |
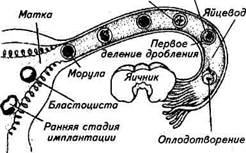
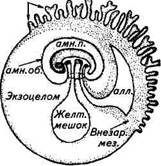
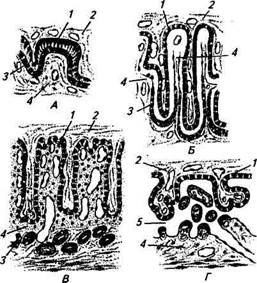
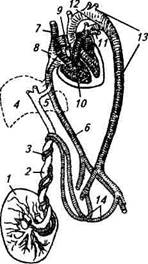 Рис. 9.8. Схема кровообращения плода млекопитающих:
Рис. 9.8. Схема кровообращения плода млекопитающих: