|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Значення спинного мозку в регуляцiї тонусу м'язiв. Гама-петляСкладнi рухи можуть бути здiйсненi лише при умові, що до ефекторних iмпульсів постiйно вносити- муться поправки з урахуванням тих змiн, якi вiдбуваються кожну мить у м'язi в процесі його скорочення. Тому м'язова система є джерелом численної аферентної iмпульсацiї. Спинний мозок постiйно одержує iнформацiю про ступiнь напруження м'язових волокон та їх довжину. Рецепторна частина аналiзатора руху являє собою м'язовi веретена й сухожильнi органи Гольджi. М'язовi веретена. У м'язах, здебільшого розгиначах, що виконують антигравітаційну функцію, є м'язовi во- локна, котрi тоншi й коротшi за iншi. Вони розмiщуються невеликими пучками (вiд 2 до 12 волокон) у сполучнотканиннiй капсулi. Через форму подiбнi структури одержали назву м’язових веретен (рис. 4.8).
А – сегментарна локалізація патологіч- ного вогнища: 1 – спинно-таламічний шлях (провідник больової і температур- ної чутливості), 2 – пірамідний шлях (забезпечує довільні рухи), 3 – тонкий пучок (провідник суглобно-м’язового й тактильного відчуттів); В – схема локалі- зацій порушень чутливостей і рухів. М’язовi волокна, що розмiщенi в капсулi, названi iнтрафузальними (лат. fusus – веретено), тодi як зви- чайнi волокна, на частку яких припа- дає головна маса м'язу, названi ек- страфузальними, або робочими волокнами. Веретено одним кiнцем прикрiплюється до перимiзiю екст- рафузального м'язового волокна, другим – до сухожилля. Центральна частина iнтрафузального волокна є власне рецепторною частиною. Iснує два типи iнтрафузальних м’язових волокон, якi вiдрiзняються за розмiщенням ядер: ядра волокон з ядерним ланцюгом i ядра волокон з ядерною сумкою. Очевидно, цi два типи волокон функцiонально відрізняються.
Рис. 4.8. Схема будови м’язового вере- тена (за Р. Шмидт, Г.Тевс, 1985)
Рис. 4.9. Схема здійснення міотатичного рефлекса. Сухожильнi органи Гольджi – особливi рецептори, якi складаються з сухожильних ниток, що вiдходять приблизно вiд 10 екстрафузальних м’язо-вих волокон i фiксуються до сухожиль м'за послідовно, у виляді ланцюга. Адекватним подразником для них є зміна напруги м’яза. До органiв Гольджi підходять товстi мiєлiновi волокна групи Ib (Аb). У сухожильному органі вони розгалужуються на тонші чисельні гілочки й втрачають мієлін. Такі рецептори розповсюджені в скелетних м’язах. Характер збудження м'язових веретен i сухожильних органiв залежить вiд їх розмiщення: м’язовi веретена з'єднуються паралельно, а сухожильнi органи – послiдовно вiдносно екстрафузальних м’язових волокон. Отже, як наслідок, м’язовi веретена сприймають головним чином довжину м'яза, а сухожильнi органи – його напругу. Чутливi закiнчення м’язових веретен можуть збуджуватись не тiльки пiд впливом розтягування м'яза, але й у результатi скорочення iнтрафузальних м’язових волокон при збудженнi γ-мотонейронiв. Цей механiзм носить назву γ-петлi (рис. 4.10). При скороченнi тiльки iнтрафузальних волокон довжина або напруга м'яза не змiнюється, однак при цьому розтягується центральна частина цих волокон i тому збуд- жуються чутливi закiнчення. Таким чином, iснує два механiзми збудження м’язових веретен: 1) розтягування м'яза i 2) скорочення iнтрафузальних волокон; цi два механiзми можуть дiяти синергiчно. Дата добавления: 2015-12-15 | Просмотры: 928 | Нарушение авторских прав |
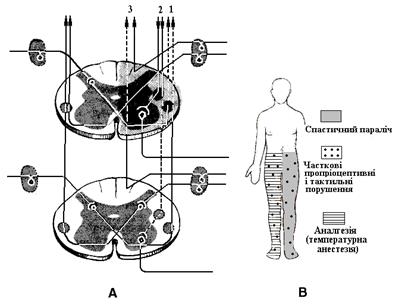 Рис. 4.7. Синдром пошкодження половини поперечного перерізу спинного мозку (синдром Броун- Секара).
Рис. 4.7. Синдром пошкодження половини поперечного перерізу спинного мозку (синдром Броун- Секара). Аферентна iннервацiя. У кожне веретено проникає товсте мiєлiнове нервове волокно; воно посилає гiлочку до кожного iнтрафузального волокна й закiнчується на його середнiй частинi, спіралеподібно оплітає її та створює так звані аннуло- спіральні закінчення. Цi аференти є волокнами Iа (Аa), а їх закiнчення носять назву первинних чутливих закiнчень. Адек- ватним подразником для них є зміна та швидкість зміни довжини м'язового волокна (рис. 4.9). Частина веретен iннервується аферентними волокнами групи II (Аb). Цi чутливi волокна «об- слуговують» виключно iнтрафузальнi волокна з ядерним ланцю- гомі мають назву вторинних сенсорних закінчень; розташову- ються вони своїми відростками периферично від анулоспіраль- них закінчень. Їх збудливість нижча, а чутливість до динамічних параметрів менша.
Аферентна iннервацiя. У кожне веретено проникає товсте мiєлiнове нервове волокно; воно посилає гiлочку до кожного iнтрафузального волокна й закiнчується на його середнiй частинi, спіралеподібно оплітає її та створює так звані аннуло- спіральні закінчення. Цi аференти є волокнами Iа (Аa), а їх закiнчення носять назву первинних чутливих закiнчень. Адек- ватним подразником для них є зміна та швидкість зміни довжини м'язового волокна (рис. 4.9). Частина веретен iннервується аферентними волокнами групи II (Аb). Цi чутливi волокна «об- слуговують» виключно iнтрафузальнi волокна з ядерним ланцю- гомі мають назву вторинних сенсорних закінчень; розташову- ються вони своїми відростками периферично від анулоспіраль- них закінчень. Їх збудливість нижча, а чутливість до динамічних параметрів менша. Еферентна iннервацiя iнтрафузальних м’язових во- локон здiйснюється нервовими волокнами групи А- g. Нер- вова клiтина, вiд якої вони вiдходять, є γ-мотонейроном.
Еферентна iннервацiя iнтрафузальних м’язових во- локон здiйснюється нервовими волокнами групи А- g. Нер- вова клiтина, вiд якої вони вiдходять, є γ-мотонейроном.