|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Функцiї заднього мозкуЗаднiй мозок – це найбiльш древнiй вiддiл головного мозку, який ще зберiгає деякi риси сегментарної
Функцiї заднього мозку (як i спинного) подiляють на двi групи: 1) провiдниковi, 2) рефлекторнi. Провiдниковi функцiї по- в'язанi з низхiдними й висхiдними провiдниковими шляхами спинного мозку, а також зi шляхами, що закiнчуються (кортикобуль- барний) i починаються (оли- воспинальний, вестибулос- пинальний, ретикулоспи- нальний) у довгастому моз- ку.
Рис. 4.17. Ядра черепних нервів (дорзальна проекція). Чутливі ядра зображено чорним кольором, рухові – сірим.
Рефлекторнi функцiї заднього мозку зумовленi дiяльнiстю черепномозкових нервiв (V-VII – мiст, VIII-XII – довгастий мозок). Виключно руховими є VI, XI, XII пари. Решта ядер змiшанi: в їх складi є руховi, чутливi й вегетативнi нейрони. Головними вегетативними структурами довгастого мозку є ядернi утворен- ня X пари (n.vagus), у меншiй мiрi VII i IX пар, а також ядернi утворення ретикулярної формацiї (РФ) (гiганто-целюлярне, дрiбноклiтинне й латеральне ядра).
У довгастому мозку розташовано наступні рефлекторнi центри. 1. Бульбарний дихальний центр, який складається з двох пiвцентрiв – центру вдиху та центру видиху. Бульбарний ди- хальний центр утворений за участю n.tr.solitarius, n.ambiguus та ядер ретикулярної формації. У верхній частині мосту розташо- ваний пневмотаксичний центр, який регулює змiну актiв вдиху й видиху, тобто частоту дихання. Існує уявлення, що в нижній частині мосту знаходиться апнейстичний центр (регулятор глибини дихання). 2. Центр серцево-судинної дiяльностi справляє вплив на роботу серця (за участю IX, X пар черепних нервiв), а також на тонус кровоносних судин. Нейрони цього центру перебувають весь час у станi тонiчного збудження за рахунок iмпульсiв, якi надходять рiзними аферентними шляхами, а також завдяки хемочутливостi. В умовах клінічної патології (наприклад, при порушенні лікворотоку) подразником може бути й підвищений внутрішньочерепний тиск. Цi два центри є життєво-необхiдними. 3. Травний центр (його бульбарна частина) забезпечує первинну переробку їжi шляхом жування (V), смоктання (V, VII, IX), ковтання (V, VII, IX, X), слиновидiлення (VII, IX); регулює моторику шлунка та ки- шечника (X), секрецiю шлункового соку, соку пiдшлункової залози, активність секреторних клітин печінки та видiлення жовчi (X, РФ). 4. Центри захисних рефлексiв: мигання (III), чихання (IX, X, XI), секреції бронхіальних залоз, кашлю (IX, X, XI), iкання (n.phrenicus), блювання (X). 5. Центри рухових рефлексiв, якi є аналогами сегментарних спинальних (пов'язанi з ядрами VII, IX, XI, XII пар черепних нервiв), забезпечують скорочення мiмiчних м'язiв, м'язiв гортанi, шиї, потилицi; у новона- роджених – хоботковий рефлекс. Більшість цих рефлексів значущі для фонації та артикуляції. 6. Бульбарний центр м'язового тонусу структурно представлений латеральним ядром вестибулярної частини VIII пари (ядро Дейтерса), а також ядрами РФ (рис. 4.19). Вiд ядра Дейтерса починається вести- булоспинальний тракт, неперехрещений, прямує до переднiх рогiв спинного мозку, закiнчується моноси- наптично або через вставнi нейрони на a- i g-мотонейронах екстензорiв. У РФ стовбура мозку видiляють двi частини. Вiд мосту починається неперехрещений медiальний рети- кулярний тракт. Вiд довгастого мозку починається латеральний ретикулоспинальний тракт; цей тракт складається як з перехрещених, так i неперехрещених волокон. Обидва тракти закiнчуються в сiрiй речовинi спинного мозку достатньо далеко вiд моторних ядер. Волокна вiд РФ довгастого мозку збуджу- ють a - i g-мотонейрони згиначiв i гальмують мотонейрони розгиначiв, тодi як волокна вiд мосту мають протилежну дiю.
Функцiональне призначення довгастого мозку в регуляцiї м’язового тонусу можна виявити в експе- риментах з децеребрацiєю. Поперечна перерiзка стовбура мозку мiж довгастим i середнiм мозком виконується iнтерколiкулярно – з таким розрахун- ком місця перетину, щоб червонi ядра середнього мозку залишилися вище лiнiї перерiзки (головний мозок видаляється). Виникає стан децеребрацiйної ригiдностi. Полягає цей стан у рiзкому пiдвищеннi тонусу (гiпертонусi) м'язiв-розгиначiв кiнцiвок, тулуба, шиї. Характерною буде поза
тварини: переднi й заднi кiнцiвки, хвiст випрямленi, тулуб вигнуто дугою донизу, голова закинута назад. Стан децеребраційної ригідності може виникнути й у людини при пошкодженнях середнього мозку (рис. 4.20). Причина пiдвищення тонусу екстензорiв пов'язана з переважаючим активуючим впливом вестибу- лоспинального тракту (збережені зв’язки від ядра Дейтерса) i виключенням впливу центрiв середнього мозку (червоного та ретикулярних ядер) (рис. 4.21). Ригiднiсть зникає у децереброваної тварини пiсля перерiзання заднiх корiнцiв спинного мозку. Пов'язано це з тим, що ригiднiсть зумовлена γ-еферентною петлею за рахунок надходження iмпульсiв вiд g-мотонейронiв по g-волокнах до iнтрафузальних м’язових волокон м'язiв екстензорiв. Iнтрафузальнi волокна скорочуються, збуджуються рецептори ядерної сумки; iмпульси чутливими Iа (Аa) волокнами у складi заднiх корiнцiв надходять у спинний мозок до a- мотонейронiв, активують їх, що призводить до скорочення екстензорiв. Така ригiднiсть зветься g- ригiднiстю. Таким чином, децеребрацiйна ригiднiсть має рефлекторну природу й обумовлена збуджен- ням рецепторів вестибулярного апарату, пропрiорецепторiв м'язiв шиї i пропрiорецепторiв великих розгиначiв.
рефлекси звуться шийними рефлексами. Рис. 4.20. Децеребраційна ригідність. Децереброваним тваринам властивi тiльки статичні рефлекси. Реалiзуються такi рефлекси в нерухомому станi при змiнах пози й тому звуться рефлексами пози. Якщо тонус змiнюється завдяки збудженню вес- тибулярного апарату, такi рефлекси звуться лабiринтними тонiчними рефлексами. У разi збудження рецепторiв м'язiв шиї, такi позно-тонiчнi
Рис. 4.21. Механізми розвитку децеребраційної ригідності. При цьому вiдбувається змiна тонусу екстензорiв усiх кiнцiвок: пiдвищення (при повертаннi голови вгору на 45˚ вiдносно горизонтальної площини), або зни- ження (при нахилi на 45˚ донизу). Шийнi рефлекси вивчаються при зруйнуваннi лабiринтiв на децереброваних тваринах (рис. 4.22). Якщо після такого втручання голову тварини вiдкинути назад, тодi рефлекторно пiдвищується тонус розгиначiв переднiх кiнцiвок i знижується тонус розгиначiв заднiх кiнцiвок. При нахиленнi голови дони- зу пiдвищується тонус розгиначiв заднiх кiнцiвок i знижується тонус розгиначiв переднiх кiнцiвок. При поворотi голови праворуч пiдвищується тонус екстензорiв правих кiнцiвок; при поворотi лiворуч – тонус екстензорiв лiвих кiнцiвок. Це є проявом можли- востей грубого перерозподілу тонусу між м’язами кінцівок. До статичних рефлексiв, що здiйснюються у децереброваної тварини, належить також компенса- торне установлення очей. При повертаннi голови вгору очні яблука опускаються донизу і, навпаки, при опусканні голови – доверху. Це теж є проявом перерозподілу тонусу. Його біологічне значення полягає у фіксації предмета в полі зору, але лише нерухомого. Захисне значення мають згинальні шкіряні реф- лекси. Вони полісинаптичні. Викликаються подразнен-
ням рецепторів шкіри (больових, температурних та ін.). Призначення таких – уникнути пошкоджуючого подразника.
А. Пасивне підіймання голови (вказано стрілкою) супроводжується зменшенням тонусу розгиначів задніх кінцівок і підвищенням тонусу розгиначів передніх кінцівок. Б. Пасивне нахилення голови (вказано стрілкою) викликає протилежний ефект.
Отже, якщо спинний мозок лише готує органі- зацію протидії силі тяжіння на рівні окремих м’язів- розгиначів за механізмом g-петлі, то задній мозок реалізує комплексне підвищення тонусу екстензо- рів тулуба і кінцівок як передумову антигравітації. Та за участю довгастого мозку ефективної антигравітації ще немає, оскільки відсутня рівновага. Якщо децеребро- вану тварину поставити на випрямлені кінцівки, то поза “стояння” підтримуватиметься лише при умові, що центр ваги буде проектуватися на площину опори, або якщо притулити такий препарат до стінки, тобто при наявності опори. Значне підвищення тонусу м’язів шиї (подібно до стану децеребраційної ригідності) у людини виникає під впливом високого внутрішньочерепного тиску, тобто при безпосереднь- ому механічному подразненні бульбарного центру м’язового тонусу. Елементи ригідності розгиначів спостерігаються при деяких хореографічних та спортивних вправах (“спад назад” на перекладині, стійка балерини “на одному пальці”, стійка спортсмена донизу головою на руках). Дата добавления: 2015-12-15 | Просмотры: 921 | Нарушение авторских прав |
 будови первинної мозкової трубки. Проявом сегментарностi є розташу- вання в ньому 8 пар череп- них нервiв (V-XII пари) (рис. 4.17), а також забезпечення сегментарних рефлексiв. Ядра черепних нервiв дея- кою мiрою гомологiчнi цен- трам спинномозкових метамерiв; вони пов'язанi з рiзними черепномозковими центрами й низхiдними шля- хами спинного мозку (рис. 4.18).
будови первинної мозкової трубки. Проявом сегментарностi є розташу- вання в ньому 8 пар череп- них нервiв (V-XII пари) (рис. 4.17), а також забезпечення сегментарних рефлексiв. Ядра черепних нервiв дея- кою мiрою гомологiчнi цен- трам спинномозкових метамерiв; вони пов'язанi з рiзними черепномозковими центрами й низхiдними шля- хами спинного мозку (рис. 4.18). Рис. 4.18. Шкірна іннервація голови та шиї гілками трійчас- того нерва та нервами, які походять із верхніх шийних сегментів спинного мозку.
Рис. 4.18. Шкірна іннервація голови та шиї гілками трійчас- того нерва та нервами, які походять із верхніх шийних сегментів спинного мозку. Рис. 4.19. Схема розташування рухових центрів у стовбурі мозку – у довгастому мозку, мосту та середньому мозку. У разі порушення зв’язків головно- го мозку з відділами, які розташовані нижче трьох ліній перерізки, отримують децеребровану тварину (1), мезен- цефальну тварину (2), і спинальну тварину з високою перерізкою (3).
Рис. 4.19. Схема розташування рухових центрів у стовбурі мозку – у довгастому мозку, мосту та середньому мозку. У разі порушення зв’язків головно- го мозку з відділами, які розташовані нижче трьох ліній перерізки, отримують децеребровану тварину (1), мезен- цефальну тварину (2), і спинальну тварину з високою перерізкою (3).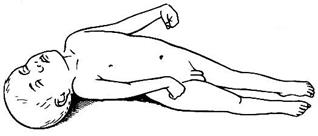
 Лабiринтнi рефлекси на децереброваних тваринах вивчаються при фiксуваннi голови до тулуба за до- помогою гiпсової пов'язки, або пiсля перетину заднiх шийних корiнцiв. Тварину перемiщують у просторi.
Лабiринтнi рефлекси на децереброваних тваринах вивчаються при фiксуваннi голови до тулуба за до- помогою гiпсової пов'язки, або пiсля перетину заднiх шийних корiнцiв. Тварину перемiщують у просторi.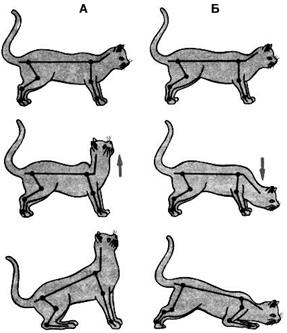 Рис. 4.22. Позні рефлекси у децереброваної тварини з видаленими лабіринтами.
Рис. 4.22. Позні рефлекси у децереброваної тварини з видаленими лабіринтами.