|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Базальні ядра (nuclei basales)
До базальних ядер (БЯ) належать: 1) хвостате ядро, n.caudatus 2) лушпина, putamen 3) блiда куля, globus pallidus 4) чорна речовина, substantia nigra 5) субталамiчна дiлянка, regio subthalamica 6) огорожа, claustrum 7) мигдалеподiбне тiло, corpus amygdaloideum Хвостате ядро i лушпину віднесено до neostriatum, а блiду кулю – до paleostriatum. Neostriatum i paleostriatum разом звуться смугастим тiлом, corpus striatum. БЯ - додаткова до пірамідної кортикоспинальної система регуляції моторних функцій (стара її назва – екстрапірамідна система). Як і мозочок, БЯ не мають прямого зв’язку з мотонейронами й впливають на них через інші рухові центри. БЯ отримують аферентнi зв'язки вiд усiх дiлянок кори великих пiвкуль, особливо від асоціативних і мо- торних; вiд таламусу й чорної субстанції (дофамiнергiчний шлях); еферентнi зв'язки вiд стрiатума більшістю через блiду кулю прямують до чорної речовини, таламусу (вентрального переднього й вен- трального латерального ядер) i до покрiвлi середнього мозку. Є прямі зв’язки з моторною корою (рис.
Рис. 4.26. Найважливіші аферентні, еферентні й внутрішні зв’язки базальних ядер. СЦ – серединний центр; ПВЯ – переднєвентральне ядро; ВЛЯ – вентрола- теральне ядро; СЯ – субталамічне ядро.
БЯ вiдiграють головним чином роль промiжної ланки в ланцюговi, який з'єднує моторнi дiлянки кори з рештою її дiлянок. У цьому вiдношеннi БЯ подiбнi до мозочка: БЯ, як і пiвкулi мозочка із зубчастими ядрами, можна вважати аферентними по вiдношенню до прецентральної моторної дiлянки кори; як і мозочок, посилають сигнали до кори через ядра таламусу. Головними функціями БЯ є регуляція м’язового тонусу із значною перевагою гальмівних впливів; регуляція рухів; організація й реалізація рухових програм та інших поведінкових реакцій. Все це здійснюється з певною своєрідністю, зі значною питомою вагою гальмівних впливів (БЯ вважаються «гальмівним ситом» для кори мозку). Головним призначенням БЯ є організація контро-
лю комплексних патернів рухів, здебільшого поведінкових, із впливом на інтенсивність, напрямок, послідовність різноманітних наступних і паралельних рухів для досягнення специфічних моторних наслідків, тобто певної мети. Отже, БЯ забезпечують загальний руховий фон, збагачують головний рух допоміжними, співдружніми (розмахування руками під час ходьби, бігу, стрибків; жестикуляція при розмові). Механізм та призначення полягає в підвищенні рівня активності робочих центрів реципрокним шляхом, що забезпечує високу ефективність головного рухового акту. БЯ регулюють скорочення дрібних м’язових груп і рухи в дрібних суглобах, забезпечуючи цілеспрямованість, влучність рухових актів. Саме втручанням БЯ забезпечується емоційне забарвлення обличчя (тобто міміки) та інтонації голосу (мови). Завдяки регуляції м’язового тонусу за пластичним типом (на його подолання потрібне безперервне зу- силля) забезпечується злитність, плавність та економічність рухів. Участь БЯ у здійсненні рухових програм стосується впливів на їх формування, а також здійснення; на ініціацію та завершення, особливо для повільних рухових компонентів; нарешті – на запам’ятовування. За останніми даними БЯ належить важлива роль у переході від задуму (фази підготовки) до обраної програми дії (фази виконання). Серед рухових програм, організованих за участю БЯ, є як генетично успадковані, так і придбані за умовнорефлек- торним принципом тренування. Структурною основою для таких є петлі з залученням певних ядер стріарної системи, якими циркулює збудження. Наприклад, разом з моторними ділянками кори цикл лушпини забезпечує складні рухи, що потребують попереднього навчання (письмо, гра на музичному інструменті та ін.). Цикл розпочинається в премо- торних і додаткових моторних ділянках кори, поширюється в напрямку лушпини до внутрішньої частини блідої кулі (повз хвостаті ядра), потім до таламусу й знов до кори мозку. Є петлі за участю стріарних структур, які спонукають до їжі, пізнавальної діяльності, статевої поведінки та ін. Се- ред петель нижчого порядку – окуломоторна: від поля 8 фронтальної кори й 7 поля тім’яної – через хвостате ядро, бліду кулю, чорну речовину, верхні горбики чотиригорб’я. Ця петля контролює спрямовані рухи очей при загальмовуванні саккад. Окремим структурам стріатуму властиве певне функціональне приурочення. Так, лушпині властиве спонукання до їжової поведінки. Її подразнення викликає слиновиділення, зміни дихання, пошкодження спричиняє трофічні розлади шкіри й внутрішніх органів (гепато-лентикулярну дегенерацію). Бліда куля провокує їжову поведінку (жування, ковтан- ня), орієнтовні реакції. Огорожа пов’язана з цибулиною й нюховою корою, причетна до організації орієнтовних реакцій на сенсорні стимули. Її руйнування призводить до втрати можливості розмовляти. Хвостате ядро, роль якого виявляється головним чином у пригніченні палеостріатуму, а також кори й підкірки, гальмує безумовну та умовнорефлекторну поведінку. При пошкодженні неостріатуму мають місце розлади орієнтування у просторі, порушення пам’яті, щезання умовних рефлексів на тривалий термін, і утруднення їх вироб- лення. У людини, під час нейрохірургічних операцій, стимуляція хвостатого ядра порушує мовний контакт. Серед медіаторів стріарної системи крім ДОФА (від чорної речовини) мають значення ацетилхолін і глутамат (від волокон кори), ГАМК (від хвостатих ядер), серотонін (від ядер шва), норадреналін (від голубої плями), а також ряд пептидів – тобто ціла низка синаптоактивних речовин. Саме тому замісна терапія a-ДОФА не забезпечує повноцінного відновлення функцій при патологічних ураженнях стріарної системи. Поширеною патологічною моделлю порушень БЯ є синдром Паркiнсона (тремтливий паралiч). Хво- рих з цiєю патологiєю легко впiзнати за маскоподiбним обличчям, вiдсутністю або рiзким зменшенням жестикуляцiї, обережною ходою мiлкими кроками й тремтiнням. При неврологiчному дослiдженнi у цих хворих виявляються такi симптоми, як акiнезiя, ригiднiсть, постійне тремтiння, яке зникає лише уві сні, хробакоподібні рухи (атетоз). Суть акiнезiї полягає в тому, що хворий вiдчуває великi, а iнколи непереборнi труднощi при початку й завершеннi рухiв. Ригiднiсть являє собою постiйне збiльшення м'язового тонусу за типом пластичної або воскоподiбної ригiдностi, незалежно від того, у стані спокою чи руху перебувають суглоби. При пасивних рухах розслаблення ступінчасте (симптом зубчастого колеса) Тремтiння спокою бiльше проявляється в дистальних вiддiлах кiнцiвок, зникає пiд час цiлеспрямованих рухiв i вiдновлюється пiсля їх закiнчення. У дрібних суглобах розвиваються хробакоподібні рухи. З патофiзiологiчної точки зору акiнезiю слід розглядати як порушення програмування рухiв. Синдром Паркінсона пов’зують головним чином з дефіцитом у постачанні дофаміну компактною частиною чорної речовини й, отже, порушенням у зв’язку з цим нігро-стріатних зв’язків. Саме недостатність дофаміну спричиняє надмірне збудження (розгальмовування) нейронів хвостатого ядра й підсилений його гальмівний вплив на палеостріарні структури. Та використанням для замінної терапії α-ДОФА (препарату дофаміну, здатного проникати через гемато-енцефалічний бар’єр) повністю проявів паркінсонізму не ліквідується. Мабуть крім дофаміну мають значення й інші медіатори – глутамат, ацетилхолін, пептиди тощо. Проявом надмірної активності палеостріарних структур є гіперкінетичний синдром, пов’язаний з ураженням неостріарних структур (хвостатого ядра) і «перезбудженням» нижче розташованих (зокрема, блідої кулі). Характер- ним є надмірність рухової активності у вигляді постійно відновлюваних, неритмічних крупнорозмашистих рухів, у тому числі й подібних до свідомих (непосидючість, гримаси, смішливість, неартикульована фонація, емоційна нестриманість). М’язовий тонус знижений з переважанням екстензорного. Уві сні ці розлади зникають. Підсумовуючи значення БЯ в системі надсегментарних структур мозку, слід відзначити своєрідність співідношень цього ядерного комплексу з корою як найвищої підкоркової надбудо- ви. Стріато-коркові зв’язки, опосередковані через таламус, особливо розвинуті з Дата добавления: 2015-12-15 | Просмотры: 742 | Нарушение авторских прав |
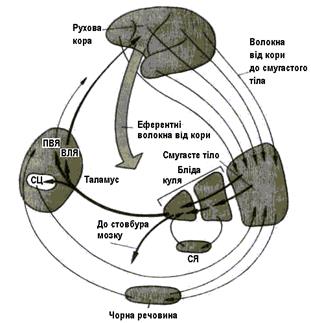 4.26).
4.26).