|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Тонка кишка (порожня, дванадцятипала)У тонкій кишці здійснюються процеси подальшого травлення кор-му під дією ферментів власних та позастінних травних залоз і всмок-тування поживних речовин корму в кровоносне та лімфатичне русло. Оптимальними умовами дії ферментативних систем тонкої кишки є слаболужне середовище. Кишковий сік містить різні ферменти, які діють на білки та полі-пептиди (трипсин, ентерокіназа, кіназоген, пептидази), нуклеопротеї-ди (нуклеаза), вуглеводи (амілаза, мальтаза, сахароза, лактоза), жири (ліпаза). Ферментативна система посмугованої облямівки епітеліаль-них клітин забезпечує пристінкове (примембранне) перетравлюван-ня корму. Клітини дисоційованої ендокринної системи забезпечують Спеціальна гістологія
регулящю секрецп кишкового соку, кровопостачання і відпо-відну інтенсивність процесів всмоктування та моторики тон-кої кишки. Кишечник розвивається із кількох ембріональних зачатків. Це ентодерма — епітеліальна ви-стилка кишки, мезенхіма — спо-лучнотканинні та м'язові струк-тури, вісцеральна несегментова-на мезодерма — мезотелій. Стінка порожнього кишеч-ника, як типовий трубкоподіб-ний орган, складається з слизо-вої, м 'язової та серозної оболонок (рис. 152). Особливістю будови стінки кишечника є наявність по-стійних структур, здатних значно збільшувати всмоктувальну по-верхню епітеліального шару сли-зової оболонки. До цих структур відносять кишечні складки (від-повідають циркулярним у люди-ни), у формуванні яких беруть участь усі шари слизової обо-лонки. Кишкові ворсинки — це інтердигітації основног пластин-ки, покриті епітеліальним шаром; крипти (ліберкюнові залози) — трубкоподібні впинання епітелію у власну пластинку, посмугована облямівка — плазмолема апікаль-ного полюса циліндричних енте-роцитів, яка утворює мікровор-синки (останні не слід змішувати із ворсинками тонкої кишки).
Рис. 152. Будова стінки порож-ньої кишки: 1-слизова оболонка; 2-м'язова оболон-ка; 3-серозна оболонка; 4-одношаровий епітелій ворсинки; 5-основна плас-тинка слизової оболонки; 6-ворсинки; 7-крипти; 8-м'язова пластинка; 9-підслизова основа; 10-кровоносні су-дини; 11-підслизове нервове сплетення (Мейснерове); 12-кільцевий шар; 13-поздовжній шар; 14-міжм'язове нервове сплетення (Ауербахове); 15-мезотелій.
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія Кишкові ворсинки, основою яких є сполучна тканина власної плас-тинки та поодинокі гладенькі міоцити — це пальцеподібні вирости сли-зової оболонки у просвіті кишки, які надають 'ш оксамитового вигляду. Епітеліальний шар слизової оболонки ворсинок утворений цилінд-ричним епітелієм, до складу якого входять три різновиди клітин: енте-роцити (стовпчасті епітеліоцити ворсинок), келихоподібні та кишкові ендокриноцити. Основними клітинами епітеліального шару є ентероцити із посмуго-ваною облямівкою. Ці клітини мають чітко виражену полярність: ядро знаходиться у базальній частині ентероцита, а на апікальному полюсі розміщуються мікроворсинки, які під світловим мікроскопом мають ха-рактерний вигляд посмугованої облямівки. Висота мікроворсинки досягає близько 1 мкм, вони складаються із інтердигітацій плазмолеми ентероцита, їх добре видно в електронному мікроскопі. Кожна мікроворсинка має субмікроскопічний каналець, що не лише збільшує всмоктувальну поверхню, а й надає їй властивостей пористого каталізатора. Каймисті ентероцити мають овальне ядро, до-бре розвинену ергастоплазму, лізосомний апарат. У цитоплазмі, під кай-мою розміщується центросома, над ядром — комплекс Гольджі. Завдяки наявності як ворсинок, так і мікроворсинок всмоктувальна поверхня слизової оболонки збільшується у сотні разів. Ферментативна активність в посмугованій облямівці значно вища, ніж у просвіті кишки, через це процеси розщеплення та всмоктування тут відбуваються зна-чно інтенсивніше, ніж у порожнині кишечника. Апікальна частина енте-роцита містить тонофіламенти, з участю яких формуються замикальні пластинки та щільні контакти, що закривають міжклітинні щілини й стають непроникними для речовин з просвіту кишки. На поверхні мікроворсинок знаходиться глікокалікс (суміш гліко-протеїдів). Він міцно зв'язаний із плазмолемою і забезпечує адсорбцію речовин на поверхні ентероцитів. Гістохімічними методами в мікровор-синках виявляються глікопротеїдні комплекси з ферментами, які пере-шкоджають проникненню в клітину мікробів. Таким чином облямівкові ентероцити є основними функціональними елементами процесів пєре-травлювання і всмоктування у тонкій кишці. Продукти розщеплення білків і вуглеводів — амінокислоти та моносахариди транспортуються через ентероцит від апікального до базального полюса і через базальну мембрану у гемомікроциркуляторне русло сполучнотканинної основи Розділ 4 Спеціальна гістологія ворсинки. Таким шляхом засвоюється вода та розчинені в ній міне-ральні солі і вітаміни. Жири засвоюються двома шляхами: фагоцитозу крапельок емульго-ваного жиру облямівковими ентероцитами; всмоктування гліцерину та жирних кислот із наступним синтезом нейтрального жиру в цитоплазмі клітин. Ліпіди через базальний полюс каймистих ентероцитів потрапля-ють у лімфатичні капіляри. Келихоподібні клітини розміщуються в епітеліальному шарі між об-лямівковими ентероцитами. Це одноклітинні залози, що продукують слизовий секрет, який сприяє просуванню частинок корму до товстої кишки. У розширеній апікальній частині клітини нагромаджуються секреторні продукти, а до базального полюса відтиснуті ядро та добре розвинені агранулярна ендоплазматична сітка, комплекс Гольджі, міто-хондрії. Після виділення секрету келихоподібні клітини набувають ци-ліндричної форми. Ендокриноцити (аргирофільні клітини), так як і келихоподібні, роз-сіяні поодиноко серед облямівкових ентероцитів. їх також відносять до АРИД-системи. Апудоцити тонкої кишки синтезують біологічно ак-тивні речовини, які впливають на функцію облямівкових ентероцитів, келихоподібних клітин, гладеньких міоцитів судин стінки кишки, а звід-си — на процеси секреції, всмоктування та моторики. Основна (власна) пластинка складається із пухкої сполучної та рети-кулярної тканин, що містять значну кількість лімфоцитів, плазматичних клітин, еозинофілів. В центральній частині пластинки розміщуються лімфатичні судини, вздовж яких орієнтовані міоцити (скорочувальний елемент ворсинки), кровоносні судини та нерви. Вростання епітеліаль-ного гиару у власну пластинку утворює кишкові крипти — прості, інколи розгалужені трубчасті (ліберкюнові) залози (150-180 млн у тонкій киш-ці), які також збільшують всмоктувальну поверхню слизової оболонки. У тварин різних видів їх форма неоднакова. Серед епітеліоцитів крипт, крім клітин, охарактеризованих в складі ворсинок, є ще безоблямівкові ентероцити (стовпчасті) та екзокриноцити з ацидофільною зернистіс-тю (клітини Панета). Ендокриноцитів у кринтах більше, а функціональна активність їх така ж, як і у ворсинках. Безоблямівкові ентероцити (стовпчасті епітеліоцити) являють со-бою популяцію малодиференційованих клітин, які є джерелом фізіоло- В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія гічної регенерації епітелію крипт та ворсинок тонкої кишки. На їхньому апікальному полюсі відсутня посмугована облямівка. Поступово, у міру проліферації, диференціації і спеціалізації, клітини переміщуються від дна крипт до верхівки ворсинки. Протягом 2-3 діб епітеліоцити ворси-нок повністю змінюються. Панетовські (апікальнозернисті) клітини розміщуються на дні крипт. Вони мають призматичну форму, а на апікальному полюсі міс-тяться великі ацидофільні гранули секрету, на базальному — ядро, комп-лекс Гольджі та ендоплазматична сітка. Секрет клітин Панета містить ферменти — дипептидази, які розщеплюють дипептиди до амінокислот. Вважають, що їх ферменти також нейтралізують соляну кислоту шлун-ка, що потрапляє разом із хімусом. В основній пластинці слизової оболонки тонкої кишки також міс-тяться скупчення лімфоцитів, які утворюють підепітеліальні лімфатич-ні фолікули (солітарні фолікули) та агрегатовані пейєрові бляшки. У великої рогатої худоби вони великі, у коня та свині дрібніші. У молодих тварин і дітей їх більше, з віком їх кількість зменшується. Кишкові ворсинки — це випинання основної пластинки, покриті епі-теліальним шаром із усіма його клітинними різновидами. Вони не лише збільшують поверхню слизової оболонки, а й є основними органами всмоктування, з урахуванням особливостей гемолімфатичного русла. На верхівці ворсинки артеріальна судина продовжується безпосередньо у вену, яка потім збирає у себе всі капіляри ворсинки. Під час всмоктуван-ня кров рухається через капіляри, а в спокої — через артеріо-венозний анастомоз. Строма, судини та епітелій ворсинки з'єднані волокнами ре-тикулярної тканини і діють під час скорочення як одне ціле. Функціонує ворсинка як всмоктуючий насос за рахунок скорочення міоцитів. До-вжина ворсинок у тварин різних видів — різна — у хижаків вони довші, у травоїдних — коротші. М'язова пластинка має типову будову. Підслизова основа тонкої кишки утворена пухкою сполучною та ретикулярною тканинами. У ній міститься значна кількість кровонос-них та лімфатичних судин, нервових сплетень. Лімфатичні фолікули підслизової основи необхідно розглядати як прониклі сюди із власної пластинки підепітеліальні лімфатичні вузлики. М'язова оболонка тонкої кишки утворена двома шарами гладеньких міоцитів: внутрішнім та зовнішнім. Внутрішній шар розвинений біль-ше, його клітини розміщуються циркулярно, зовнішнього — у косопбвз- Розділ 4 Спеціальна гістологія довжньому напрямку Між шарами м'язової оболонки у пухкій сполуч-ній тканині розміщується м'язове нервове сплетення. Перистальтичні скорочення структур цієї оболонки забезпечує переміщення та про-штовхування хімусу в товсту кишку Серозна оболонка утворена пух-кою сполучною тканиною, яку покриває один шар клітин мезотелію. Слід зазначити, що окремі відділи тонкої кишки мають ряд морфо-логічних особливостей, але будову дванадцятипалої кишки і, особливо, її слизової оболонки буде описано нижче. Дванадцятипала кишка — це відділ тонкої кишки з найбільш інтен-сивним впливом хімічних сполук ферментів на корм, як за рахунок се-крету власних пристінних, так і застінних залоз (печінки, підшлункової залози). Для слизової оболонки дванадцятипалої кишки характерне утво-рення широких та коротких ворсинок, а насиченість ворсинками одини-ці площі слизової оболонки — максимальна. Крім крипт, які розміщуються в основній пластинці, в підслизовій основі дванадцятипалої кишки знаходяться кінцеві секреторні відділи складних, розгалужених, трубчастих, дуоденальних (брунерівських) за-лоз. У жуйних залози трубчасті, в інших тварин - альвеолярно-трубчасті. Кінцеві секреторні відділи дуоденальних залоз складаються з паріеталь-них ентероцитів, мукоцитів (клітин Панета), келихоподібних клітин та ендокриноцитів. Вивідні протоки доуденальних (брунерівських) залоз проникають через м'язову пластинку і відкриваються у основи крипт або між сусідніми ворсинками. Вивідні протоки утворені мукоцитами кубічної або призматичної форми, а біля поверхні слизової оболонки вони замінюються облямів-ковими ентероцитами. Секрет дуоденальних залоз покриває слизову оболонку, захищає її від шкідливого впливу кислоти шлункового соку, нейтралізуючи останню. Дипептидази та амілази брунерівських залоз розщеплюють дипептиди й вуглеводи відповідно до амінокислот та мо-ноцукрів. Інші оболонки дванадцятипалої кишки відповідно мають таку ж будову, як вже описані вище структури кишкової трубки. Товста кишка У цьому відділі кишкової трубки процеси перетравлення корму не-значні. Тут відбувається інтенсивне всмоктування води та мінеральних речовин, а бактеріальна флора — синтезує вітаміни групи ВіК,а також В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія нагромаджуються екскреторні речовини (продукти метаболізму), солі тяжких металів тощо. Особливості функції відбиваються на гістологічній будові товстих кишок (рис. 153), а саме: кишкові ворсинки є лише у зародка, у дорос-лих тварин їх немає; крипти виділяють незначну кількість ферментів і побудовані простіше. Облямгвка ентероцитів розвинена слабо; є велика кількість келихоподібних клітин, особливо в криптах; лімфатичні вуз-лики містять центри розмноження лімфоцитів.
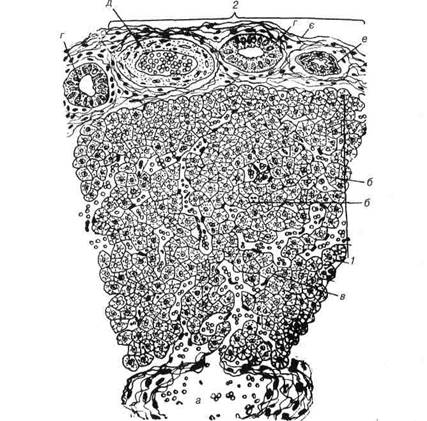
2*<
Рис. 153 Будова стінки товстої кишки: 1-слизова оболонка; 2-м'язова оболонка; 3-серозна оболонка; 4-підслизова основа; 5-одношаровий епітелій; 6-келихоподібні клітини; 7-крипти; 8-влас-на пластинка; 9-м'язова пластинка; 10-підслизове нервове вплетення (Мей-снерове); 11-лімфатичнийфолікул; 12-кровоноснісудини; 13-кільцевийшар; 14-поздовжній шар м'язової оболонки; 15-мезотелій серозної оболонки. Розділ 4 Спеціальна гістологія Отже, товстим кишкам притаманна загальнобіологічна законо-мірність будови: слизова, м'язова та серозна оболонкн складаються із шарів певних структур. Названі вище особливості будови найбільше стосуються слизової оболонки. Епітеліальний шар останньої утворений одношаровим, циліндричним облямівковим епітелієм. Випинання епі-телію в основну пластинку формує крипти. Епітеліальний шар слизо-вої оболонки і крипт містить кілька видів клітин: облямівкові ентеро-цити, келихоподібні клітини, безоблямівкові ентероцити та незначну кількість ендокриноцитів і клітин Панета. Переважну більшість клітин епітеліального пласта становлять кели-хоподібні клітини, які продукують велику кількість слизу. Біля основи крипт локалізуються безоблямівкові, стовпчасті ентероцити з високою мітотичною активністю, завдяки проліферації яких здійснюється фі-зіологічна регенерація епітелію. Прошарки пухкої сполучної тканини, що розміщуються між криптами, продовжуються в тканину основної пластинки. Остання має велику кількість ретикулярної тканини, у якій скупчуються лімфоцити, утворюючи великі лімфатичні вузлики із цен-трами розмноження. Вони можуть проникати через м'язову пластинку й зливатися з аналогічними структурами підслизової оболонки. М'язова пластинка має типову будову. В підслизовій основі у пух-кій тканині розміщуються судинне та підслизове нервове сплетення. Лімфатичні вузлики, з'єднуючись, утворюють солітарні фолікули, а останні — пейєрові бляшки. Поздовжній шар м 'язової оболонки міо-цитів утворює тенії (крім великої рогатої худоби), між якими м'язова оболонка значно тонша. Між шарами циркулярних та поздовжніх міо-цитів розміщується міжм'язове (Ауербахове) нервове сплетення. Спо-лучна тканина серозної оболонки покрита мезотелієм. Стінка прямої кишки має такі ж оболонки. Епітеліальний шар сли-зової оболонки у каудальній частині змінюється від одношарового стовпчастого до багатошарового сквамозного. В стінці прямої кишки лімфатичні вузлики досягають найбільшого розвитку. У деяких свій-ських тварин у стінці локалізуються анальні залози слизового типу, а у хижих — навколоанальні — подібні до жирових залоз. Очеревина Серозна оболонка черевної порожнини, паріетальний та вісцераль-ний листки якої вистилають стінку та внутрішні органи, виконує ряд В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія функцій: виділяє незначну кільгсість серозног рідини для зменшення тер-тя органів ггри перистальтиці, бере участь в обміні речовин та формує захисний біологічний бар 'єр. Мезотелій, що покриває очеревину, є похідним целомічної вистил-ки. Це одношаровий плоский епітелій, який здатний до всмоктування за допомогою мікроворсинок, що містяться на апікальній поверхні клі-тини і являють собою інтердигітації плазмолеми. Одночасно з цим, ме-зотелій виділяє певну кількість рідини в черевну порожнину. Мезотелій надзвичайно чутливий до різних подразнень, навіть легенький дотик до нього викликає відшарування клітин. Це явище мезотелію добре проявляється при запальних процесах, коли клітини його у великій кількості відшаровуються від очеревини, ггеретворюючись у макрофа-ги і надходять до перитонеальної рідини. Сгголучнотканинна основа очеревини має своєрідний еластично-ко-лагеновий комплекс, який забезпечує її функцію при зміні положення органів черевної порожнини. Строма очеревини має ряд різновиднос-тей у будові від 6 до 14 шарів в брижах. Очеревина має розвинену інтрамуральну нервову систему, яка складається з двох нервових сплетень: гговерхневого вузькоггетлистого та глибокого широкопетлистого. Крім того, в неї надзвичайно різнома-нітний рецеггторний апарат — від вільних арборизацій до спеціалізо-ваних інкапсульованих закінчень. Це пояснює велику тактильну чут-ливість очеревини при різних патологічних змінах внутрішніх органів. Таким чином очеревина — є своєрідним біологічним локатором, який надзвичайно чутливий до зміни середовища черевної порожнини. Печінка Печінка — найбільша застінна залоза шлунково-кишкового тракту з надзвичайно різноманітними функціями. У ній знезаражується вели-ка кількість продуктів обміну речовин, інактивуються гормони, біоген-ні аміни, а такожряд лікарських препаратів. Печінка бере участь у за-хисних реакціях організму проти мікробів та чужорідних речовин у ви-падку проникнення їх в організм. У ній синтезується глікоген-головне джерело підтримки ггостійног концентрації глюкози в крові. В печінці синтезуються найважливіші білки гглазми крові: альбуміни, ггротром-бін, фібриноген та ін. В ній метаболізується залізо і утворюється жовч, необхідна для засвоєння жирів у кишечнику. Розділ 4 Спеціальна гістологія Велике значення має печінка в обміні холестерину, який є важли-вим компонентом всіх клітинних мембран. В ній нагромаджуються необхідні для організму жиророзчинні вітаміни — А, Д, Е, К та ін. Крім того, в ембріональному періоді печінка виконує функцію органа кро-вотворення. Ембріогенез печінки починається з третього тижня розвитку зарод-ка із мішкоподібного випинання вентральної стінки середньої кишки (печінкова бухта, яка в процесі росту ділиться на краніальний і кау-дальний відділи). 3 краніального відділу утворюється печінка та пе-чінкова протока, а каудальний є джерелом розвитку жовчного міхура та жовчної протоки. Епітеліальні клітини краніального відділу бухти формують численні тяжі, між якими розміщується сітка широких кро-воносних капілярів, які разом із жовчною веною дають початок розви-тку воротній вені. Утворена таким чином залозиста паренхіма печінки своєю будо-вою нагадує губку. Подальше диференціювання печінки відбувається у другій половині внутрішньоутробного розвитку та в перші роки піс-ля народження, при цьому, в її паренхіму вростає сполучна тканина, супроводжуючи розгалуження воротної вени, ділить її на печінкові частки. Необхідно зазначити, що більшість із названих функцій печін-ки виконують одні і ті ж клітини, з яких складається паренхіма. Тому кожна клітина має зв'язок як з кровоносними судинами, так і з вивід-ними протоками. Все це і зумовлює гістологічну будову печінки зовсім не схожу на будову будь-якої залози і характеризується розміщенням кровоносних судин, жовчних проток і рядів клітин у вигляді сітки. Гістологічна будова печінки. Поверхня органа покрита сполучнот-канинною капсулою, яка щільно зростається з вісцеральним листком брюшини. Структурно-функціональними одиницями печінки є печінко-ві частки, які утворюють її паренхіму За класичним визначенням частки мають форму шестигранних призм, ширина яких не перебільшує 1,5 мм, а висота дещо більша. Зли-ваючись по дві і більше вони формують складні печінкові частки (рис. 154). Кількість часток у печінці людини досягає 500 тис. Міжчасточкова сполучна тканина утворює строму органа, яка роз-виненау різних представників ссавців по-різному Найбільша кількість міжчасточкової сполучної тканини у свиней, через це у них краще, ніж у інших тварин, виражена часточкова структура. У коней значно гір- 25 - 8-305 В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія ше помітна часточковість, ще гірше у жуйних і, особливо, у гризунів і хижаків, внаслідок того, що часточки відмежовані одна від одної лише міжчасточковими судинами та жовчними протоками. Інтенсивний розвиток сполучної тканини призводить до атрофїї (зменшення) пе-чінкових часток і є ознакою захворювання печінки на «цирроз» (від гр. кіггпоз — рудий).
Рис. 154. Гістоструктура печінки: 1-печінкова частка; а-центральна вена; б-печінкові балки; в-гепатоцити; 2-тріада: г-міжчасточкова жовчна протока; д-міжчасточкова вена; е-міжчасточкова артерія; є-пухка сполучна тканина.
Розділ 4 Спеціальна гістологія .—------------------------------------------------------------------------ —------------------------- — Печінкові частки утворені печінковими балками та внутрішньо-часточковими синусоїдними капілярами. Печінкові балки, побудовані з гепатоцитів — (від гр. пераг - печінка, сутдіз — клітина), розміщених у радіальному напрямку (рис. 155). Між ними в тому ж напрямку від периферії до центру частки проходять кровоносні капіляри (рис. 156).
Рис. 155. Схема гістоструктури печінкової частки ссавців: 1-печінкова артерія; 2- печінкова вена; 3- жовчна протока; 4- печінкова бал-ка із гепатоцитів; 5-ендотелій печінкового синусоїда: 6-центральна вена; 7-венозний синус; 8-жовчні капіляри (за Хемом) Внутрічасточкові кровоносні капіляри складаються із одного шару плоских ендотеліоцитів; в місцях з'єднання ендотеліальних клітин утворюються дрібні пори. Ці ділянки ендотелію називають ситоподібними. Між ендотеліоцитами розсіяні численні зірчасті макрофаги (клі-тини Купфера), які не утворюють суцільного шару. На відміну від ен-дотеліоцитів, вони мають моноцитарне походження і є макрофагами 25* В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія печінки, з якими пов'язані її захисні функцїї. Зірчасті макрофаги ма-ють відросчасту форму і будову, подібну до фагоцитів. При здійсненні захисних реакцій вони відходять від стінок кровоносних капілярів і перетворюються у вільні макрофаги.
Рис. 156. Ін'єкція судин печінки: 1-центральна вена; 2-внутрішньочасточкові синусоїди; 3-септальна вена; 4-зона залягання печінкових балок; 5-міжчасточкові вени. Базальна мембрана на значному проміжку стінки внутрішньодоль-кових капілярів відсутня, за винятком периферичних та центральних відділів. Навколо капілярів утворена вузенька (0,2-1 мкм) навколосинусо-їдна щілина, в яку через пори в ендотелії капілярів потрапляють скла-дові частини плазми крові, а в умовах патології проникають і формені елементи. В навколосинусоїдній щілині, крім рідини, багатої на білки, знаходяться мікроворсинки гепатоцитів, інколи відростки зірчастих Розділ 4 Спеціальна гістологія макрофагів, аргирофільні волокна, які обплітають печінкові балки, а також відростки клітин — перисинусоїдальних ліпоцитів. Ці клітини мають невеликі розміри (5-10 мкм), й розміщуються між сусідніми ге-патоцитами. У їх цитоплазмі постійно знаходяться дрібні крапельки жиру, міститься багато вільних рибосом, поодинокі мітохондрії. Кіль-кість перисинусоїдальних ліпоцитів різко збільшується при деяких хронічних захворюваннях печінки. Є дані, які показують, що ліпоцити, подібно до фібробластів, здатні до волокноутворення, а також є депо жиророзчинних вітамінів. Печінкові балки складаються з гепатоцитів, з'єднаних між собою десмосомами за типом «замка«. Балки анастомозують між собою і тому їх радіальний напрямок у частках не завжди чітко помітций. В печінкових частках гепатоцити розміщуються двома рядами, тісно контактуючи між собою, через те на поперечному зрізі кожна балка із ряду парних гепатоцитів. За аналогією з іншими залозами печінкові балки вважають кінцевими відділами печінки, оскільки утворюючі їх гепатоцити секретують глюкозу, білки крові та інші речовини. Між рядами гепатоцитів розміщуються жовчні капіляри, діаметром від 0,5 до 1 мкм, які не мають власної стінки. їх стінка утворена зіткне-ними поверхнями гепатоцитів, на поверхні яких містяться невеликі заглиблення, що збігаються один з одним і разом утворюють просвіт жовчного капіляра (рис. 157). Просвіт жовчного капіляра не сполуча-ється з міжклітинною щілиною завдяки тому, що мембрани сусідніх ге-патоцитів у цьому місці щільно прилягають одна до одної, утворюючи замикаючі пластинки. Поверхні гепатоцитів, які обмежують жовчні капіляри, мають мі-кроворсинки, що проникають у їх просвіт. На звичайних гістологічних препаратах жовчні капіляри зали-шаються непомітними і їх виявляють лише при Спеціальних методах фарбування (імпрегнація сріблом або ін'єкція капілярів забарвленою масою через жовчну протоку). На таких препаратах видно, що жовчні капіляри починають-ся сліпо на центральному кінці печінкової балки, йдуть вздовж неї, утворюючи в боки коротенькі сліпі вирости, і на периферії частки переходять у холангіоли — короткі трубочки, вузький просвіт яких обмежений 2-3 овальними клітинами. Холангіоли впадають у між-часточкові жовчні протоки. В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
Рис. 157. Ультраструктура печінкової клітини та її поєднання з кровоносними капілярами і жовчними канальцями: 1-лізосоми; 2-гранулярна ендоплазматична сітка; 3-клітини ендотелію сину-са; 4-еритроцит; 5-периваскулярний простір; 6-ліпопротеїд; 7-агранулярна ендоплазматична сітка; 8-глікоген; 9-жовчний каналець; 10-мітохондрії; 11-комплекс Гольджі; 12-пероксисома. Таким чином, жовчні капіляри розміщуються всередині печінкових балок, оскільки між балками проходять кровоносні судини. Через це кожний гепатоцит у балці має два полюси, один — біліарний, звернений в просвіт жовчного капіляру, у який клітини секретують жовч, дру-гий — васкулярний, спрямований до кровоносного внутрішньочасточ- Розділ 4 Спеціальна гістологія кового капіляра, в який клітини виділяють глюкозу, сечовину, білки та інші речовини. Між кровоносними та жовчними капілярами в жодно-му місці немає безпосереднього зв'язку, оскільки їх розділяють один від одного печінкові та ендотеліальні клітини. Тільки при деяких за-хворюваннях (паренхіматозна жовтяниця), що пов'язане з частковим ураженням та загибеллю частини гепатоцитів, жовч може безпосеред-ньо із жовчних капілярів проникати в кровоносні. У таких випадках жовч розноситься кров'ю по всьому організму при жовтяниці і надає тканинам жовтого кольору. Згідно з іншим твердженням про будову печінкових часток вони складаються із широких анастомозуючих пластин, між якими зна-ходяться кров'яні лакуни, по яких повільно циркулює кров. Стінка лакуни утворена ендотеліоцитами та зірчастими макрофагами. Нині з'явилася нова теорія про гістофункціональні одиниці печінки, відмін-на від класичних печінкових часток. Згідно з нею структурно-функці-ональною одиницею органа є, так звані, портальні печінкові частки та печінкові ацинуси. Печінкові клітини — гепатоцити становлять 60% усіх клітинних елементів органа. Вони виконують більшу частину функцій, прита-манних печінці. Гепатоцити мають неправильну, багатогранну форму, діаметр їх досягає 20-25 мкм, у багатьох (до 20%) міститься два ядра. Кількість гепатоцитів залежить від функціонального стану організ-му: вагітність, лактація, голодування значно впливають на вміст їх в печінці. Ядра гепатоцитів мають круглу форму, величина їх коливається від 7 до 16 мкм, що пояснюється наявністю у печінкових клітинах, поряд із звичайними ядрами (диплоїдними), більш значніших — по-ліплоїдних. Цитоплазма печінкових клітин забарвлюється як кислими, так і основними фарбниками, оскільки містить велику кількість РНП (рибонуклеопротеїдів), у ній є всі види органел. Гранулярна ендоплаз-матична сітка має вигляд вузеньких канальців із прикріпленими рибо-сомами. В центролобулярних клітинах вона розміщена паралельними рядами, а в периферійних — в різних напрямках. Агранулярна ендоплазматична сітка у вигляді трубочок та пухир-ців розсіяна по всій цитоплазмі або концентрується у певних її ділян-ках; гранулярна — синтезує білки крові, а агранулярна бере участь у метаболізмі вуглеводів. Крім того, ендоплазматична сітка за рахунок В.П. Новак, Ю.П. Бичков, М.Ю. Пилипешо Цитологія, гістологія, ембріологія утворення в ній ферментів здійснює дезинтосикацію шкідливих речо-вин, а також інактивує деякі гормони та лікарські препарати. Біля канальців гранулярної ендоплазматичної сітки розміщують-ся пероксисоми, з якими пов'язують обмін жирних кислот. Більшість мітохондрій гепатоцита мають округлу або овальну форму й розмір від 0,8 до 2 мкм. Зустрічаються також мітохондрії нитчастої форми, до-вжина яких досягає 7 мкм і більше. Для них характерна невелика кіль-кість крист та помірно щільна матриця. Мітохондрії рівномірно розпо-діляються по всій цитоплазмі, а їх кількість в одному гепатоциті може змінюватися. Комплекс Гольджі в період інтенсивного жовчовиділен-ня зміщується в клітині до просвіту жовчного капіляра. Навколо нього розміщуються окремі лізосоми, або, навіть, цілі групи. На васкулярній та біліарній поверхнях гепатоцитів розміщуються мікроворсинки. Печінкові клітини містять ряд включень: глікоген, ліпіди, пігменти та інші, які утворюються із поживних речовин, що приносяться кров'ю. Кількість їх змінюється у різні фази діяльності печінки, особливо зна-чні зміни спостерігаються у зв'язку з станом травлення. Через 3-5 го-дин після приймання їжі кількість глікогену в гепатоцитах збільшує-тья і досягає максимуму через 10-12 год. Після 24-48 год. глікоген пе-ретворюється у глюкозу і зникає із цитоплазми клітини. У випадках, коли корм багатий на жири, в цитоплазмі гепатоцитів з'являються кра-плинки жиру, причому раніше в клітинах, які розміщені на периферії печінкових часток. При деяких захворюваннях нагромадження жиру в клітинах може набувати патологічного стану — ожиріння, яке має міс-це при травмах мозку, променевій хворобі, алкоголізмі. Для печінки характерний добовий ритм секреторних процесів: вдень переважає виділення жовчі, вночі — синтез глікогену. Очевидно, цей ритм регулюється з участю гіпоталамуса та гіпофіза. Жовч та глі-коген синтезуються у різних зонах печінкової частки: жовч утворюєть-ся в периферійній зоні і лише потім цей процес поступово поширюєть-ся — на центральну зону, а синтез глікогену здійснюється в зворотному напрямку — від центра до периферїї частки. Гепатоцити виділяють в кров глюкозу, сечовину, білки, жири, а в жовчні капіляри — жовч. Кровообіг у печінці. Виходячи із класичних уявлень про будову печінкових часток, кровоносну систему печінки умовно поділяють на три частини: систему притоку крові до часток, систему циркуляції крові всередині них, систему відтоку крові від часток. Розділ 4 Спеціальна гістологія Систему притоку представляє воротна вена та печінкова артерія. В печінці вони багаторазово поділяються на дрібніші судини: часточкові, сегментарні, міжчасточкові та навколочасточкові вени та артерії. Ра-зом судини воротної вени, печінкової артерії та жовчні протоки фор-мують тріади. Поруч з ними розміщуються лімфатичні судини. Між-часточкові артерії і вени розміщуються на боковій поверхні часток, а навколочасточкові судини, які відгалужуються від них, обплітають часточки на різних рівнях. Міжчасточкові та навколочасточкові вени мають слабо розвине-ну м'язову оболонку, але в місцях розгалуження формують сфінктери. Відповідні часточкові та навкочасточкові артерії відносять до судин м'язового типу Від названих судин беруть початок кровоносні капі-ляри, які входять у печінкові частки й, зливаючись, утворюють вну-трішньочасточкові синусоїдні судини, що формують систему цирку-ляції крові у печінкових часток. По цих судинах змішана кров тече в напрямку від периферїї до центру часток. Співвідношення між венозною та артеріальною кров'ю у внутріш-ньочасточкових судинах регулюється станом сфінктерів міжчасточ-кових вен. Внутрішньочасточкові капіляри відносять до синусоїдного типу капілярів. У них базальна мембрана несуцільна. Вони розміщу-ються між тяжами гепатоцитів — печінковими балками, радіально схо-дячись до центральних вен, що лежать у центрі печінкових часток. Система відтоку крові починається центральними часточковими венами, які при виході із часток впадають у збиральні, підчасточкові вени, що проходять в міжчасточкових перетинках. Підчасточкові вени не супроводжуються артеріями та жовчними протоками, оскільки вони не входять до складу тріад. За цією ознакою їх легко відрізнити від су-дин системи воротної вени — міжчасточкових та навколочасточкових вен, що приносять кров до часток. Центральні та підчасточкові вени — судини безм'язового типу. Зливаючись, вони утворюють початок печінкових вен, які виходять із печінки і вливаються в каудальну порожнисту вену Сфінктери, які розміщуються на початку печінкових вен, регулюють відток крові від часток і всього органа залежно від її хімічного складу та маси. Таким чином, печінка забезпечується кров'ю від воротної вени та печінкової артерії, завдяки чому через печінку за незначний проміжок часу проходить вся кров організму, збагачуючись білками та звільню- В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія ючись від метаболічних речовин азотистого обміну, від їх шкідливих домішок. Паренхіма печінки має велику кількість кровоносних капі-лярів, у результаті чого кровоток у печінкових часточках відбувається повільніше, що сприяє обміну між кров'ю та клітинами печінки, які ви-конують захисну, знезаражуючу, синтетичну та інші важливі для орга-нізму функції. При необхідності в судинах печінки може депонуватися велика кількість крові. Жовчовивідні шляхи. До них відносять внутрішньопечінкові та зо-внішньопечінкові жовчні протоки. Стінка міжчасткових жовчних про-токів, в які збираються із печінкових часток жовчні капіляри, склада-ється із одношарового кубічного епітелію. У міру збільшення діаметра, клітини епітелію стають вищими і набувають призматичної форми. У великих протоках з'являється м'язовий шар із кільцево розміщених гладеньких м'язових клітин. Таким чином, між частками артерія, вена та жовчний капіляр роз-міщуються разом, утворюючи печінкову тріаду. Жовчний міхур. Стінка його складається із трьох оболонок: сли-зової, м 'язової та адвентиції, а з боку черевної порожнини покрита очеревиною. Слизова оболонка утворює численні складки, покрита високими призматичними епітеліальними клітинами, які мають об-лямівку, а у жуйних містяться ще й келихоподібні епітеліоцити. В ді-лянці шийки міхура розміщуються альвеолярно-трубчасті залози, які виділяють слиз. Епітелій слизової оболонки має здатність всмоктува-ти з жовчі воду та деякі інші речовини. В підепітеліальній пластинці розміщуються лімфатичні вузлики. М'язова оболонка утворена пучка-ми гладеньких міоцитів циркулярного напрямку, що в ділянці шийки утворюють сфінктер. Адвентиція — типова щільна сполучна тканина, в якій є велика кількість еластичних волокон, що утворюють сіточки. Іннервація печінки здійснюється вегетативним нервовим сплетен-ням, яке розміщується під капсулою органа. Нервові волокна за ха-рактером субстрату, який вони іннервують, поділяють на секреторні, вазомоторні та чутливі. Секреторні нервові волокна всередині часток утворюють закінчення на поверхні печінкових балок, а деякі між гепатоцитами. Вазомоторні — це гілочки симпатичного нерва, які відіграють велику роль у регуляції циркуляції крові та наповнення печінки як депонуючого органа. Чутли-ві нервові волокна знаходяться в міжчастковій сполучній тканині, на су- Розділ 4 Спеціальна гістологія динах та всередині часток, де формують арборизації. Для судин печінки характерна багата чутлива іннервація і, особливо, вен воротної системи. По ходу нервових стовбурів розміщуються окремі нервові клітини. Тканина печінки здатна до високої фізіологічної та репаративної регенерації. Процеси регенерації відбуваються шляхом мітотичного поділу гепатоцитів, а також компенсаторним збільшенням їх розмірів (гіпертрофії). У деяких тварин до 50% тканини печінки регенерує за 10-15 днів. 3 віком ця пластичність знижується, у гепатоцитах збільшу-ється кількість ліпофусцину, ядра гіпертрофуються за рахунок збіль-шення вмісту ДНК, збільшується також кількість сполучної тканини між печінковими частками. Дата добавления: 2015-12-15 | Просмотры: 1498 | Нарушение авторских прав |