|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Моногибридное скре-щивание. Закон единооб-разия гибридов первого поколения. Доминант-ность и рецессивност. Кодоминантность. Ал-лельное состояние гена.Моногибридное скрещивание рассмотрим на примере красноцветкового гороха с белоцветковым. Во время мейоза у гибридного растения F1 материнские хромосомы несущие доминантный ген, и отцовский хромосом несущий рецессивный ген расходятся в дочерней клетке независимо др. от др., и поэтому при случайном соединении гамет во время оплодотворения образуется три типа зигот. Красноцветковые растения F2 одинаковы по фенотипу, но различны по генотипу. Организмы содержащие в соматических клетках 2 доминантных или 2 рецессивных гена данной аллельной пары называються гомозиготными, а организмы содержащие разные гены данной аллельной пары – гетерозиготными. Гомозиготные особи при размножении не дают расщепление, а гетерозиготные продолжают расщепляться. Правило единообразия гибридов первого поколения. При опылении красноцветкового гороха пыльцой, взятой с растений с белыми цветками, все гибриды первого поколения имели красную окраску цветков Такие же результаты были получены при обратном скрещивании, когда белоцветковые растения опылялись пыльцой красноцветковых. Следовательно, все гибридные растения первого поколения имели одинаковую красную окраску цветков, т. е. были по этому признаку единообразны. Единообразие гибридов первого поколения наблюдалось Г. Менделем во всех скрещиваниях, которые он проводил. Это дало ему основание сформу-лировать одну из основных закономерностей наследования — правило единообразия гибридов первого поколения. Явление доминирования. В примере, взятом из опытов. Г. Менделя по скрещиванию растений гороха с разноокрашенными цветками, признаки красной и белой окраски, составляющие одну пару, проявлялись у потомства по-разному. Красная окраска цветков у гибридов неизменно сохранялась, белая подавлялась и не обнаруживалась. Признак, проявляющийся у гибридов первого поколения, в данном случае красная окраска цветков, Г. Мендель назвал доминантным, а не проявляющийся, в данном опыте белая окраска цветков,— рецессивным. Подавление у гибридных организмов одних признаков другими получило в генетике название доминирования. В потомстве могут одновременно проявляться признаки обоих родителей. Этот тип наследования получил название кодоминирования. Его примером может служить наследование групп крови у человека. Если один из родителей имеет группу крови А, а другой Б, то в крови детей присутствуют антигены характерные как для группы А так и для группы Б. Гены одной пары признаков находятся в одинаковых точках гомологичных хромосом. Такие гены получили название аллельных. Понятие аллельности — одно из важнейших. В генетике оно имеет такое же значение, как понятие валентности в химии. Явления наследственности могут быть поняты и объяснены только на основании представления об аллельности дискретных наследственных единиц (генов). Материальной основой распределения аллельных генов при образовании гамет является процесс мейоза.
14 Гаплоидия, противоположное полиплоидии явление, заключающееся в кратном уменьшении числа хромосом у потомства в сравнении с материнской особью. Г., как правило, — результат развития зародыша из редуцированных (гаплоидных) гамет или из функционально равноценных им клеток путём апомиксиса, т. е. без оплодотворения. Г. редко встречается в животном мире, но распространена у цветковых растений: зарегистрирована более чем у 150 видов растений из 70 родов 33 семейств (в т. ч. из семейства злаков, паслёновых, орхидных, бобовых и др.). Г. генетически детерминирована и встречается у некоторых видов и сортов с определённой частотой (например, у кукурузы — 1 гаплоид на 1000 диплоидных растений). В эволюции видов Г. служит своеобразным механизмом, снижающим уровень плоидности. Г. пользуются для решения ряда генетических проблем: выявления эффекта дозы гена, получения анеуплоидов, для исследования генетики количественных признаков, генемного анализа и др. В селекции растений Г. пользуются для получения из гаплоидов путём удвоения у них числа хромосом гомозиготных линий, равноценных самоопылённым линиям при производстве гибридных семян (например, у кукурузы), а также для перевода селекционного процесса с полиплоидного на диплоидный уровень (например, у картофеля). Особая форма Г. — андрогенез, при котором ядро спермия замещает ядро яйцеклетки, используется для получения мужских стерильных аналогов у кукурузы. В соответствии с этой классификацией выделяют следующие группы гаплоидных растений: Классификация: Матроклинные гаплоиды – растения, произошедшие от яйцеклетки с редуцированным числом хромосом, или из клеток зародышевого мешка выполняющих функции яйцеклетки. К этому типу относят подавляющее большинство гаплоидов. Андрогенные гаплоиды – гаплоидные растения, развивающиеся из яйцеклетки или клеток зародышевого мешка, хромосомы которых замещены хромосомами спермия. Этот вид гаплоидии известен у небольшого числа видов. Андроклинные гаплоиды – гаплоидные растения, произошедшие из клеток мужского гаметофита – пыльцевых зерен. Получение андроклинных гаплоидов возможно только экспериментальным путём. Гаплоиды могут возникать спонтанно, но частота их спонтанного возникновения очень мала. Искусственным путем с Использованием методов in vitro удается получить большие количества гаплоидных растений. Существует три способа получения гаплоидов с использованием метода культуры изолированных тканей: андрогенез – получение гаплоидных растений на искусственной питательной среде из изолированных пыльников и микроспор. гиногенез – получение гаплоидных растений на искусственной питательной среде из изолированных семяпочек; партеногенез – получение гаплоидов из гибридного зародыша, у которого из-за несовместимости хромосом родителей потеряны отцовские хромосомы.
15. ДНК основной материальный носитель наследственности. Исследования, установившие роль аминокислот в наследственности. Почти вся ДНК находится в хромосомах — структурах клеточного ядра, являющихся материальными носителями наследственности организмов. В различных организмах содержится разное количество ДНК. Но у одного и того же организма в различных клетках (их ядрах) ее количество одинаково, хотя сами клетки значительно отличаются друг от друга по химическому составу. Мутагенное действие различного рода излучений и химических веществ на организмы связано в первую очередь с изменением ДНК. Прямым доказательством генетической роли ДНК служат опыты по бактериальной трансформации. В опытах Ф. Гриффитса мыши, которым вводили вирулентный штамм, погибали. При введении невирулентного штамма они оставались живыми. Клетки вирулентного штамма, предварительно убитые нагреванием, также не вызывали заболевания. Но результаты были получены у четвертой группы мышей, которым вводили смесь невирулентных и вирулентных, но убитых нагреванием клеток. Эти мыши заболевали инфекционной пневмонией и тоже погибали, как и мыши первой группы, которым вводили вирулентный штамм. В выделениях таких больных животных обнаруживались капсульные вирулентные клетки пневмококков. Следовательно, взаимодействие невирулентных и убитых нагреванием вирулентных клеток восстанавливало свойства и внешние признаки последних. Происходила трансформация — передача особенностей одних клеток другим Самое интересное в этих опытах заключалось в том, что трансформация происходила под влиянием какого-то вещества небелкового характера, поскольку клетки донора предварительно были убиты. в 1944 г. в экспериментах американских микробиологов-генетиков под руководством О. Эвери было показано, что этим веществом, обладающим высокой трансформирующей активностью, является чистая ДНК. Очень наглядно и точно генетическая роль ДНК была установлена А. Херши и М. Чейз благодаря использованию изотопной метки при изучении размножения фага Т2. Белок фага был помечен радиоактивной серой (35S), а ДНК — радиоактивным фосфором (32Р).Такой препарат фага смешивали с суспензией бактериальных клеток. После этого в потомстве фага с помощью специальных счетчиков радиоактивности прослеживали распределение метки. Оказалось, что новые фаговые частицы содержали только испускавший р-излучение радиоактивный фосфор, которым была помечена ДНК. Меченый белок родительского фага дочернему поколению не передавался. 35S ни у одной частицы в белковой оболочке не содержалась. Трансдукция. Поражая бактерию, ф|аг не всегда ее уничтожает. Иногда процесс вирусной инфекции протекает иначе У кишечной палочки наряду со штаммом, способным благодаря гену 1ас+ сбраживать лактозу, имеется мутантный штамм, у которого ген lac останавливает этот процесс. Если фаг, выращенный на штамме 1ас+, перенести в среду, где развивается штамм 1ас~, то некоторая часть бактерий благодаря трансдукции в результате генетической рекомбинации перейдет в форму 1ас+. Так, в опытах по трансдукции была подтверждена генетическая роль ДНК. Трансдукци-я используется для изучения структуры хромосом и тонкого строения гена, а также в экспериментах по генной инженерии. Таким образом, совокупность всех полученных в описанных исследованиях данных убедительно показывает, что ДНК — это химическое вещество, в котором организм сохраняет свои наследственные свойства, т. е. наследственная информация организма записана в структуре молекул ДНК.
16.автополиплоиды -орг-мы,получ-ся в рез-те крат-ного увелич гаплоидного набора хромосом одного и того же вида.м/быть спон-танной(обнаружена де фри-зом) и индивид-й(сорта оз.ржи 28 а не 14хром)клевер луго-вой,гречиха,корнеплоды,плодовые,ягодные.по сравне-нию с диплоидами более плодови-ты.1особенности:морфолог и анатомич увеличение размеров раст и орга-нов;2физиолог особенно-сти-замедление клеточного деления,увел периода веге-тации,низкое асмотич дав-ление,пониж усталость к факторам среды;3семенная продуктивность пониж-я из за нарушения мейо-за;наслед-е призна-ков,каждый ген представлен в 4х дозах и при самоопылении в f2 потомство дает 5 геноти-пов:1.квадроплекс все гены АААА 2.триплекс-АААа 3.дуплекс-ААаа 4.симплекс-Аааа5.нулеплекс-аааа;ращипление 35:,у диплоидов3:1;ращипление по 2 парам аллелей у дип-лоидов9:3:3:1,а автотетрап-лоидов1225:35:35:1.у авто-тетраплоидов в F2ослед поколениях сохр-ся более высокий уровень гетерози-готности,чем у диплои-дов,это м/быть исп-но для преодоления гетерозиса у гибридов в нескольких поколенияхпри переводе их из диплоидного состояния в тетра.5нескрещив-ть с дип-лоидами.изолированы не-скрещ-тью но малый % возможен(при выращивании ди- и тетра- сортов необх пространственная изоля-ция.6.исп автопо-липл.,т.к.крупные размеры цветков раст,соцветия исп в декорат цветов-ве.
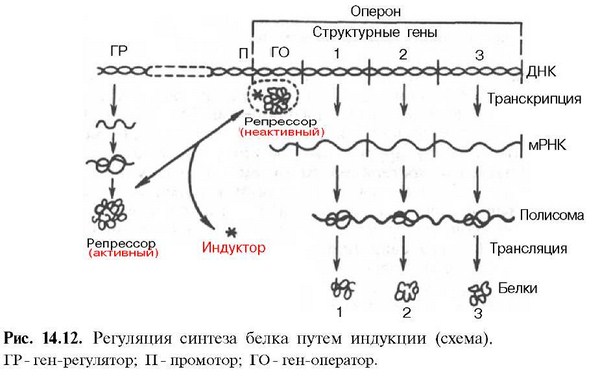
17.Регуляция синтеза белка путём индукции и репрессии. Бактериальная клетка способна запустить или прекратить синтез того или иного фермента в зависимости от присутствия соответствующего субстрата. Для этого бактериальные гены объединены в группы (кластеры) таким образом, что все ферменты, необходимые для осуществления определенного пути биосинтеза, детерминируются генами, сцепленными друг с другом. Вся группа генов может транскрибироваться в одну полицистронную м -РНК, которая последовательно транслируется рибосомами с образованием каждого из белков. Такая форма организации позволяет координировано регулировать выражение всех генов одной единицы транскрипции. Экспрессия генов упрокариот регулируется главным образом на уровне транскрипции. Роль сигнальных веществ для запуска транскрипции играют молекулы-эффекторы, представляющие собой низкомолекулярные соединения, которые являются либо субстратом для фермента, либо продуктом ферментативной деятельности соответственно. Индукция и репрессия представляют собой разные стороны одного и того же явления. Малые молекулы, индуцирующие образование ферментов, способных метаболизировать их, называются индукторами. Те же, которые предотвращают образование ферментов, способных синтезировать их, - корепрессорами. Молекулы-эффекторы не могут вступать в прямое взаимодействие с ДНК, посредником для них служит специальный регуляторный белок. Регуляторный белок, который связывается с ДНК в отсутствии индуктора, называется репрессором. За синтез регуляторных белков ответственны регуляторные гены. В присутствии белка-репрессора транскрипция блокирована; его удаление обусловливает доступ РНК-полимеразы к генам и запуск транскрипции. Прекращение синтеза фермента при помощи белка-репрессора получило название репрессии. Репрессия позволяет бактериальной клетке избежать перевода своих ресурсов на ненужную в данный момент синтетическую активность. Если индуктор присутствует в клетке в высокой концентрации, то в результате специфического присоединения к регуляторному белку он изменяет его конформацию и тем самым - его способность связываться с ДНК. Контроль транскрипции достигается взаимодействием регуляторного белка с регуляторным сайтом, называемым оператором, который расположен между структурными генами и промотором (участком, распознаваемым ДНК-зависимой РНК-полимеразой). Промотор служит местом связывания РНК-полимеразы, и от него начинается транскрипция. Совокупность промотора, оператора и структурных генов образует оперон. Оперон является функциональной генетической единицей, регулирующей экспрессию одного или группы генов Схема регуляции:
Строение оперона. Оперон - это блок генов, участвующих в обеспечении транскрипции генов, ответственных за синтез определенного генопродукта. Схема оперона: Регуляторная часть оперона: А - активатор, часть промотора, к которому присоединяется белок-активатор (САР - белок или catabolite activator protein), что активирует присоединение РНК- полимеразы к промотору; это "положительно" контролирующий элемент, который есть не в каждом опероне. П - ген-промотор - это участок ДНК, который распознается ферментом РНК - полимеразой и указывает место, где должна начинаться транскрипция. О - ген-оператор, управляющей работой структурных генов; "негативно" контролирующий элемент - присутствие на нем белка-репрессора прекращает транскрипцию. Т - ген-терминатор - это участок, после которого прекращается транскрипция и перед которым прекращается трансляция. В состав этого участка входит один из трех кодонов терминаторов (стоп-кодонов). В некоторых оперонах между оператором и структурными генами расположен участок(16 пар оснований), частью которого является аттенуатор, служащий барьером для транскрипции.
Дата добавления: 2015-12-16 | Просмотры: 1222 | Нарушение авторских прав |