|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Генетические взаимодействие между вирусами (комплементация, рекомбинация). Негенетическое взаимодействие вирусов (интерференция, фенотипическое смешение).Заражение вирусами чувствительных к ним клеток носит множественный характер, то есть в клетку может проникнуть несколько вирионов, обычно идентичных или близкородственных. В подобных ситуациях геномы вирусных частиц в динамике репродуктивных циклов могут взаимодействовать или интерферировать. Независимо от типа нуклеиновой кислоты генетические взаимодействия между вирусами представлены несколькими формами: рекомбинация, обмена фрагментами генома, комплементация. Генетические взаимодействия между вирусами. Рекомбинации и перераспределение генов вирусами Рекомбинации и перераспределение генов между геномами приводят к перераспределении генетического материала в дочерних популяциях. Они отмечены во всех группах ДНК-содержащих вирусов, у всех РНК-содержащих вирусов с сегментированным геномом и лишь у немногих РНК-содержащих вирусов с несегментированным геномом (например, у полиовируса и вируса ящура).
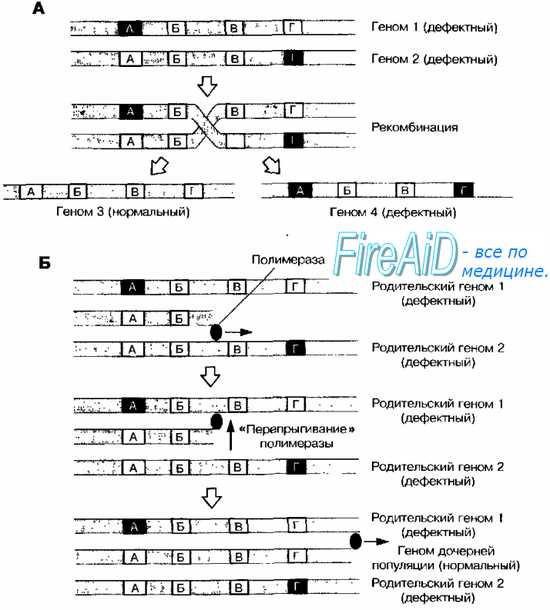
Рис. 5-5. Рекомбинация ДНК- (А) и РНК-содержащих (Б) вирусов • У ДНК-содержащих вирусов с дефектными геномами можно наблюдать рекомбинации, приводящие к образованию нормального дочернего генома (рис. 5-5, А). • У РНК-содержащих вирусов при копировании плюс-цепи в минус-цепь полимераза может переключаться с одной плюс-цепи на другую, образуя гибридную минус-матрицу РНК (рис. 5-5, Б). Подобный механизм вызывает появление генетического непостоянства у ВИЧ. Геном ВИЧ образован +РНК, поэтому при транскрипции ДНК из РНК риск «переключения» обратной транскриптазы с одной цепочки на другую достаточно велик. Если обе молекулы идентичны, то подобное явление не приводит к последствиям, но при наличии двух вирусов-мутантов в клетке возможно появление рекомбинантов с иными геномами.
Обмен фрагментами генома вирусами. Антигенный шифт
Рис. 5-6. Обмен фрагментами генома (А- Г) у РНК-содержащих вирусов Обмен фрагментами генома наблюдают у РНК-содержащих вирусов с сегментированным геномом. Б отличие от рекомбинации суть процесса состоит в обмене крупных блоков наследственного материала. Например, при множественном заражении клетки мутантами вируса гриппа с изменениями, закреплёнными в различных сегментах генома, возможно появление нормального штамма вируса; в образовании популяции последнего принимают участие геномы обоих вирусов (рис. 5-6). В частности, передача двух фрагментов генома вирулентного для цыплят штамма вируса гриппа авирулентному влечёт за собой приобретение последним вирулентных свойств. В результате обмена вирус гриппа типа А может получить новые и селективно ценные типы поверхностных Аг гемагглютинина и нейраминидазы, что обеспечивает антигенный шифт [от англ. shift, перемещение] в динамике инфекционного процесса.
Интерференция - форма негенетического взаимодействия Помимо явлений негенетической реактивации и комплементации, когда два вируса, размножаясь в одной клетке, активируют один другого, могут существовать и обратные соотношения. При смешанной инфекции клеток происходит угнетение, интерференция одного или двух взаимодействующих возбудителей. Интерференция как одна из форм негенетического взаимодействия широко известна у группы арбовирусов. Отмеченные формы взаимоотношений, встречающиеся при смешанной инфекции клеток, следует иметь в виду и отличать от истинно генетического взаимодействия между вирусами. Таким образом, к настоящему времени уже накоплен обширный материал, с достаточной убедительностью показывающий возможность широкой изменчивости арбовирусов. Это происходит как в естественных условиях циркуляции, так и при различных экспериментальных воздействиях: · длительных пассажах в организме восприимчивых и невосприимчивых животных, куриных эмбрионов, в разнообразных тканевых культурах, · под влиянием температурных условий, · а также химических и физических мутагенных факторов. Изменчивость касалась самых разнообразных наследственных признаков вирусов: · вирулентности, · иммуногенной активности, · культуральных и физико-химических свойств. Успехи исследований по аттенуации возбудителей лошадиного энцефаломиелита, Денге, клещевого и японского энцефалита значительно приблизили решение проблемы создания живых вакцин против этих инфекций. Трудности, с которыми связано получение стабильных штаммов арбовирусов, делают еще более актуальной проблему изучения изменчивости их как в экспериментальных, так и в естественных условиях с целью получения мутантов, пригодных для создания профилактических и диагностических вирусных препаратов.
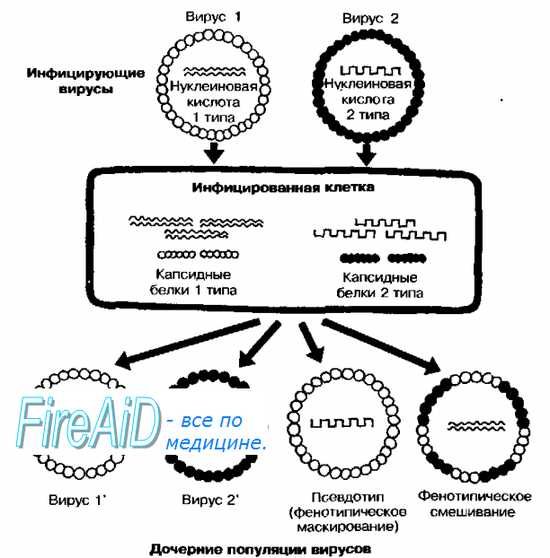
Фенотипическое смешивание и маскирование (псевдотипирование). Эти формы взаимодействий можно назвать генетическими лишь в контексте взаимодействия геномов, так как приобретаемые признаки обычно не закрепляются в потомстве, которому свойственны серологические свойства одного из «родительских* штаммов.
Рис. 5-8. Принцип образования псевдотипов (фенотипическое маскирование) и фенотипическое смешивание у вирусов. Фенотипическое смешивание вирусов наблюдают при одновременном заражении клетки близкородственными вирусами (например, различными сероварамиполиовирусов или вирусов Коксаки). В результате образуются вирионы с гибридными капсидами, в состав которых входят капсомеры, кодируемые геномами двух вирусов (рис. 5-8). Образование псевдотипов происходит при множественном инфицировании. Феномен заключается в образовании нуклеокапсида, состоящего из генома одного вируса и капсида близкородственного вируса. Генетические процессы, приводящие к образованию псевдотипов, известны как фенотипическое маскирование. Процесс может развиваться и в обратном направлении при коинфицировании вирусами идентичногопсевдотипа. Если все вирионы попавшие в клетку, содержат геном типа 2 и заключены в капсид типа 1, то дочерние популяции будут включать капсид и геном типа 2, так как образование всех их структурных компонентов кодирует геном типа 2.
Дата добавления: 2015-12-16 | Просмотры: 1275 | Нарушение авторских прав |