|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Разные постсинаптические рецепторы: ионотропное и метаботропное управлениеПредставление о рецепторах сформулировал ещё в конце XIX века знаменитый германский учёный Пауль Эрлих (Erlich P.): " Химические субстанции влияют только на те элементы ткани, с которыми они могут связаться. Эта связь должна быть специфичной, т.е. химические группы должны соответствовать друг другу, как ключ и замок". Постсинаптические рецепторы представляют собой трансмембранные белки, у которых наружная часть узнаёт и связывает молекулы медиатора. Вместе с тем, их можно рассматривать ещё и как эффекторы, управляющие открытием и закрытием хемозависимых ионных каналов. Есть два принципиально отличающихся способа управления каналами: ионотропный и метаботропный. При ионотропном управлении рецептор и канал представляют собой единую макромолекулу. Если к рецептору присоединяется медиатор, то конформация всей молекулы изменяется так, что в центре канала образуется пора и через неё проходят ионы. При метаботропном управлении рецепторы не связаны с каналом напрямую и поэтому присоединение медиатора и открытие канала разделены несколькими промежуточными этапами, в которых участвуют вторичные посредники. Первичным посредником является сам медиатор, который при метаботропном управлении присоединяется к рецептору, действующему на несколько молекул G-белка, который представляет собой длинную извитую аминокислотную цепь, пронизывающие клеточную мембрану семью своими петлями. Известно около дюжины разновидностей G-белков, все они связаны с нуклеотидом гуанозинтрифосфатом (ГТФ). Присоединение нейротрансмиттера к рецептору вызывает сразу в нескольких связанных с ним молекулах G-белка, превращение бедного энергией предшественника – гуанозиндифосфата (ГДФ) в ГТФ.
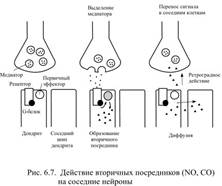
Активированный G-белок действует на интегральный белок клеточной мембраны – аденилатциклазу, которая является ферментом. Активированная аденилатциклаза вызывает превращение молекул аденозинтрифосфата (АТФ) в циклический аденозинмонофосфат (цАМФ), причём одна молекула аденилатциклазы вызывает образование множества молекул цАМФ. Молекулы цАМФ могут свободно диффундировать в цитоплазме, становясь, таким образом, переносчиками полученного сигнала внутри клетки. Там они находят ферменты – цАМФ-зависимые протеинкиназы и активирует их. Протеинкиназы стимулируют определённые биохимические реакции – характер обменных процессов направленно изменяется. Следует обратить внимание на усиление слабого синаптического сигнала при такой последовательности событий. Присоединение одной молекулы нейротрансмиттера к рецептору сопровождается активацией нескольких молекул G-белков. Каждая молекула G-белка может активировать несколько молекул аденилатциклазы. Каждая молекула аденилатциклазы вызывает образование множества молекул цАМФ. По такому же принципу, но с участием других типов G-белка активируются другие системы известных вторичных посредников (рис. 6.6). Некоторые вторичные посредники могут диффундировать через мембрану клетки и оказывать действие на соседние нейроны, в том числе и на пресинаптический (рис. 6.7). Таким, образом, ионотропное управление является непосредственным: лишь только медиатор присоединится к рецептору – открывается ионный канал, причём всё происходит очень быстро, в течение тысячных долей секунды. При метаботропном управлении ответ на присоединение медиатора непрямой, он требует участия преобразующих белков и включает активацию вторичных посредников, а поэтому и появляется значительно позже, чем ионотропный: спустя секунды, а иногда и минуты. Зато при метаботропном управлении обусловленные действием медиатора изменения сохраняются дольше, чем при ионотропном управлении. Ионотропным управлением чаще пользуются низкомолекулярные медиаторы, а нейропептиды чаще активируют системы вторичных посредников, однако эти различия не абсолютны. К ионотропным рецепторам относятся Н-холинорецепторы, один тип рецепторов для ГАМК, два типа рецепторов для глутамата, рецепторы глицина и серотонина. К метаботропным принадлежат рецепторы нейропептидов, М-холинорецепторы, альфа- и бета-адренорецепторы, по одному типу рецепторов для ГАМК, глутамата и серотонина, а также обонятельные рецепторы.
Дата добавления: 2015-12-15 | Просмотры: 1052 | Нарушение авторских прав |
 Такого рода преобразования, обусловленные присоединением остатка фосфорной кислоты, называются фосфорилированием. Вновь образующаяся связь богата энергией, поэтому молекулы G-белка, в которых произошло превращение ГДФ в ГТФ, становятся активированными (Рис. 6.4). Активация белковых молекул может проявляться в изменении их конформации, а у ферментов она обнаруживается в повышении сродства к субстрату, на который действует фермент.
Такого рода преобразования, обусловленные присоединением остатка фосфорной кислоты, называются фосфорилированием. Вновь образующаяся связь богата энергией, поэтому молекулы G-белка, в которых произошло превращение ГДФ в ГТФ, становятся активированными (Рис. 6.4). Активация белковых молекул может проявляться в изменении их конформации, а у ферментов она обнаруживается в повышении сродства к субстрату, на который действует фермент.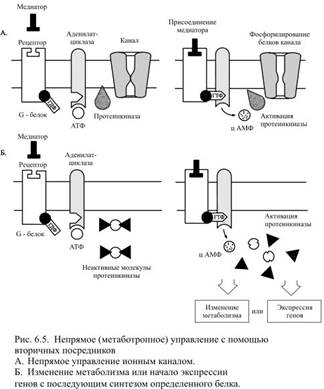 Приобретённая активность у G-белков направлена на стимуляцию или подавление активности (в зависимости от типа G-белка) некоторых ферментов (аденилатциклазы, гуанилатциклазы, фосфолипаз А 2 и С), которые в случае активации вызывают образование вторичных посредников. Конкретный ход дальнейших событий зависит от типа преобразующего сигнал белка. В случае прямого управления ионными каналами активированная молекула G-белка перемещается по внутренней поверхности мембраны к ближайшему ионному каналу и присоединяется к нему, что приводит к открытию этого канала. При непрямом управлении активированный G-белок использует одну из систем вторичных посредников, которые либо управляют ионными каналами, либо изменяют характер метаболизма – обменных процессов в клетке, либо вызывают экспрессию определённых генов, за которой следует синтез новых белков, что, в конечном счёте, тоже приводит к изменению характера обменных процессов. Из вторичных посредников лучше всего изучен циклический аденозинмонофосфат (цАМФ), образование которого осуществляется в несколько этапов (рис. 6.5).
Приобретённая активность у G-белков направлена на стимуляцию или подавление активности (в зависимости от типа G-белка) некоторых ферментов (аденилатциклазы, гуанилатциклазы, фосфолипаз А 2 и С), которые в случае активации вызывают образование вторичных посредников. Конкретный ход дальнейших событий зависит от типа преобразующего сигнал белка. В случае прямого управления ионными каналами активированная молекула G-белка перемещается по внутренней поверхности мембраны к ближайшему ионному каналу и присоединяется к нему, что приводит к открытию этого канала. При непрямом управлении активированный G-белок использует одну из систем вторичных посредников, которые либо управляют ионными каналами, либо изменяют характер метаболизма – обменных процессов в клетке, либо вызывают экспрессию определённых генов, за которой следует синтез новых белков, что, в конечном счёте, тоже приводит к изменению характера обменных процессов. Из вторичных посредников лучше всего изучен циклический аденозинмонофосфат (цАМФ), образование которого осуществляется в несколько этапов (рис. 6.5).
 Ещё один вид рецепторов находится не на постсинаптической, а на пресинаптической мембране – это ауторецепторы. Они связаны с G-белком пресинаптической мембраны, их функция состоит в регуляции количества молекул медиатора в синаптической щели. Одни ауторецепторы связываются с медиатором, если его концентрация становится чрезмерной, другие – если недостаточной. После этого меняется интенсивность выделения медиатора из пресинаптического окончания: уменьшается в первом случае и увеличивается – во втором. Ауторецепторы являются важным звеном обратной связи, с помощью которой регулируется стабильность синаптической передачи.
Ещё один вид рецепторов находится не на постсинаптической, а на пресинаптической мембране – это ауторецепторы. Они связаны с G-белком пресинаптической мембраны, их функция состоит в регуляции количества молекул медиатора в синаптической щели. Одни ауторецепторы связываются с медиатором, если его концентрация становится чрезмерной, другие – если недостаточной. После этого меняется интенсивность выделения медиатора из пресинаптического окончания: уменьшается в первом случае и увеличивается – во втором. Ауторецепторы являются важным звеном обратной связи, с помощью которой регулируется стабильность синаптической передачи.