|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
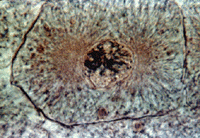
Вопрос 7. Воспроизведение клеток. Способы репродукции, их характеристика.Деление клеток Среди разновидностей клеточного деления можно назвать следующие: 1. митоз, или непрямое деление (характерное для большинства клеток); 2. амитоз, или прямое деление клеток; 3. мейоз, или редукционное деление (при образовании половых клеток); 4. эндорепродукция (образование полиплоидных и многоядерных клеток). Митоз Митоз (mitosis), кариокинез, или непрямое деление, — универсальный, широко распространенный способ деления клеток. При этом конденсированные и уже редуплицированные хромосомы переходят в компактную форму митотических хромосом, образуется веретено деления, участвующее в сегрегации и переносе хромосом (ахроматиновый митотический аппарат), происходит расхождение хромосом к противоположным полюсам клетки и деление тела клетки (цитокинез, цитотомия). Процесс непрямого деления клеток принято подразделять на несколько основных фаз: профаза, метафаза, анафаза, телофаза (рис. 20). Профаза
После окончания S-периода количество ДНК в интерфазном ядре равно 4c, так как произошло удвоение хромосомного материала. Однако морфологически регистрировать удвоение числа хромосом в этой стадии не всегда удается. Собственно хромосомы как нитевидные плотные тела начинают обнаруживаться микроскопически в начале процесса деления клетки, а именно в профазе митотического деления клетки. Если попытаться подсчитать число хромосом в профазе, то их количество будет равно 2n. Но это ложное впечатление, потому что в профазе каждая из хромосом двойная, что является результатом их редупликации в интерфазе. В профазе эти сестринские хромосомы тесно соприкасаются друг с другом, взаимно спирализуясь одна относительно другой, поэтому трудно увидеть двойственность всей структуры в целом. Позднее хромосомы в каждой такой паре начинают обособляться, раскручиваться. Двойственность хромосом в митозе наблюдается у живых клеток в конце профазы, когда видно, что общее их число в начинающей делиться клетке равно 4n. Следовательно, уже в начале профазы хромосомы состояли из двух сестринских хромосом, или, как их еще называют, хроматид. Число их (4n) в профазе точно соответствует количеству ДНК (4c). В профазе происходит конденсация хромосом. Параллельно конденсации хромосом в профазе происходят исчезновение, дезинтеграция ядрышек в результате инактивации рибосомных генов в зоне ядрышковых организаторов. Одновременно с этим в середине профазы начинается разрушение ядерной оболочки: исчезают ядерные поры, оболочка распадается сначала на фрагменты, а затем на мелкие мембранные пузырьки. Меняются в это время и структуры, связанные с синтезом белка. Происходит уменьшение количества гранулярного эндоплазматического ретикулума, он распадается на короткие цистерны и вакуоли, количество рибосом на его мембранах резко падает. Значительно (до 25%) редуцируется число полисом как на мембранах, так и в гиалоплазме, что является признаком общего падения уровня синтеза белка в делящихся клетках. Второе важнейшее событие при митозе тоже происходит во время профазы — это образование веретена деления. В профазе уже удвоенные в S-периоде центриоли начинают расходиться к противоположным концам клетки, где будут позднее формироваться полюса веретена. К каждому полюсу отходит по двойной центриоли, диплосоме. По мере расхождения диплосом начинают формироваться микротрубочки, отходящие от периферических участков одной из центриолей каждой диплосомы. Сформированный аппарат деления в животных клетках имеет веретеновидную форму и состоит из нескольких зон: двух зон центросфер с центриолями внутри них и промежуточной между ними зоны волокон веретена. Во всех этих зонах имеется большое число микротрубочек (рис. 21). Микротрубочки в центральной части аппарата деления, в собственном веретене деления, так же как микротрубочки центросфер, возникают в результате полимеризации тубулинов в зоне центриолей и около специальных структур — кинетохоров, расположенных в области центромерных перетяжек хромосом. В веретене деления принято различать два типа волокон (разной полярности): идущие от полюса к центру веретена и хромосомные, соединяющие хромосомы с одним из полюсов. В индукции роста микротрубочек веретена в зоне полюса деления принимает участие одна из центриолей диплосомы, а именно материнская. Такое новообразование и рост нитей (пучков микротрубочек) веретена происходят в профазе митоза. В то же время видны появляющиеся на хромосомах в местах первичных перетяжек (см. ниже) пластинчатые кинетохоры, около которых позднее также появляются микротрубочки, идущие в направлении полюсов деления. Таким образом, у животных клеток центриоли и хромосомные кинетохоры являются центрами организации микротрубочек веретена деления. Метафаза
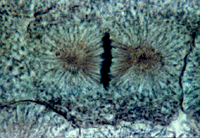
Метафаза занимает около трети времени всего митоза. Во время метафазы заканчивается образование веретена деления, а хромосомы выстраиваются в экваториальной плоскости веретена, образуя так называемую метафазную пластинку хромосом, или материнскую звезду. К концу метафазы завершается процесс обособления друг от друга сестринских хроматид. Их плечи лежат параллельно друг другу, между ними хорошо видна разделяющая их щель. Последним местом, где контакт между хроматидами сохраняется, является центромера. Анафаза
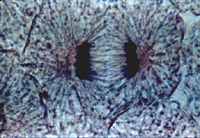
Хромосомы все одновременно теряют связь друг с другом в области центромер и синхронно начинают удаляться друг от друга по направлению к противоположным полюсам клетки. Скорость движения хромосом равномерная, она может достигать 0,2— 0,5 мкм/мин. Анафаза — самая короткая стадия митоза (несколько процентов от всего времени), но за это время происходит ряд событий. Главным из них является обособление двух идентичных наборов хромосом и перемещение их в противоположные концы клетки. Движение хромосом складывается из двух процессов, расхождения их по направлению к полюсам и дополнительного расхождения самих полюсов. Предположения о сокращении микротрубочек как о механизме расхождения хромосом в митозе не подтвердились, поэтому многие исследователи поддерживают гипотезу «скользящих нитей», согласно которой соседние микротрубочки, взаимодействуя друг с другом (например, хромосомные и полюсные) и с сократительными белками, тянут хромосомы к полюсам. Телофаза
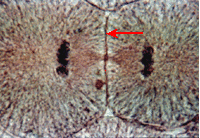
Телофаза начинается с остановки разошедшихся диплоидных (2n) наборов хромосом (ранняя телофаза) и кончается началом реконструкции новых интерфазных ядер (поздняя телофаза, ранний G1-период) и разделением исходной клетки на две дочерние (цитокинез, цитотомия). В ранней телофазе хромосомы, не меняя своей ориентации (центромерные участки — к полюсу, теломерные — к центру веретена), начинают деконденсироваться и увеличиваться в объеме. В местах их контактов с мембранными пузырьками цитоплазмы образуется новая ядерная оболочка. После замыкания ядерной оболочки начинается формирование новых ядрышек. Клетка переходит в новый G1-период. Важное событие телофазы — разделение клеточного тела, или цитотомия, или же цитокинез, - происходит у клеток животных путем образования перетяжки в результате впячивания плазматической мембраны внутрь клетки. При этом в кортикальном, подмембранном слое цитоплазмы располагаются сократимые элементы типа актиновых фибрилл, ориентированные циркулярно в зоне экватора клетки. Сокращение такого кольца приведет к впячиванию плазматической мембраны в области этого кольца, что завершается разделением клетки перетяжкой на две дочерние. При повреждении митотического аппарата (действие холода или агентов, вызывающих деполимеризацию тубулинов) может произойти или задержка митоза в метафазе, или рассеивание хромосом. При нарушениях репродукции центриолей могут возникать многополюсные и асимметричные митозы и т.д. Нарушения цитотомии приводят к появлению гигантских ядер или многоядерных клеток. Дата добавления: 2015-12-15 | Просмотры: 1258 | Нарушение авторских прав |