|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Регуляция сокращений желудка.1) Рефлекторная: а) Безусловнорефлекторная. Рецепторное поле в полости рта, пищевода, желудка, двенадцатиперстной кишке, толстой кишке. Дуга может замыкаться на различных уровнях 1. МСНС (местный уровень регуляции). 2) вегетативные ганглии; 3) на уровне нервных центров симпатической и парасимпатической систем. Эфферентное влияние: n. V. – усиливает; симпатическая система вызывает снижение моторики и угнетение эвакуации из желудка. Условнорефлекторные механизмы (на вид, запах, разговор пище). Гуморальная регуляция – гастроинтестинальные гормоны. Усиление моторики – мотилин, гистамин, серотонин Торможение – катехоламины, холецистокинин, соматостатин. 3. Дыхательный центр (понятие). Отделы ДЦ и их функции (бульбо-понтийный, спинномозговой, гипоталамический, корковый отделы). 1) Дыхательный центр. Это совокупность нейронов, обеспечивающих координацию деятельности дыхательной мускулатуры и приспособление деятельности дыхательной системы к изменившимся условиям. По последним представлениям о нервных центрах. ДЦ располагается на различных уровнях ЦНС: спинном, БПО, ЛРК, кора. Роль различных отделов в регуляции дыхания. I) Ствол мозга – здесь находится жизненно важный отдел ДЦ – бульбо – понтийный. Варолиев мост – в передней части находятся нейроны, обладающие тонической активностью, называется пневмотаксический центр. Роль: 1.обеспечивает смену дыхательных фаз (вдох – выдох); 2. увеличивает скорость развития вдоха; 3. повышает возбудимость нейронов выключающих вдох (нарушение связи). II)Продолговатый мозг. Инспираторные нейроны образуют центр вдоха. В нем можно выделить 2 отдела (воспринимающий и эффекторный). Функции инспираторных нейронов: 1) самовозбуждаются, но при условии: а) их связи с другими нейронами ДЦ, среди которых, возможно, есть пейсмекеры; б) наличии афферентных сигналов или сигналов с хеморецепторов. 2) воспринимают сигналы от хеморецепторв; 3)передают сигналы к инспираторным мышцам. Экспираторные нейроны образует центр выдоха преимущественно в вентральных ядрах. Также 2 части: воспринимающая и исполнительная. Функции экспираторных нейронов. 1) воспринимают сигналы от механорецепторов легких. 2) от проприорецепторов дыхательных мышц. 3) тормозят инспираторные нейроны, обеспечивая смену вдоха на выдох. III) Гипоталамус: 1) автоматизированное управление через АНС и ЖВС при поступлении сигналов - с интерорецепторов; - с проприорецепторов; - с терморецепторов (тепловая одышка, растет ЧД и отдача тепла). Лимбическая система изменяет дыхание при поведенческих реакциях. Кора БП: 1) тормозит ДИ; 2) условные рефлексы; 3) произвольная регуляция. 4.
Билет №8 1. 2. Переход содержимого желудка в кишечник. Пищеварение в 12ти перстной кишке. Переход содержимого желудка в двенадцатиперстную кишку, или эвакуация, вызывается чередующимися открытием и закрытием пилорического сфинктера. Механизм - когда содержимое желудка, пропитанное желудочным соком, поступает в его пилорическую часть, рецепторы в этом месте раздражаются HCl и сфинктер открывается. Часть содержимого желудка переходит в двенадцатиперстную кишку. Реакция в кишечнике становится кислой вместо щелочной, и теперь та же соляная кислота, действуя на рецепторы слизистой оболочки двенадцати перстной кишки, рефлекторно вызывает закрытие пилорического сфинктера. Когда под влиянием щелочных соков (поджелудочный и кишечный соки, желчь) соляная кислота нейтрализуется, а большая часть поступившей массы переместится дальше по кишечнику, весь процесс повторяется снова. Одна из основных функций двенадцатиперстной кишки заключается в приведении pH поступающей из желудка пищевой кашицы к щелочному, не раздражающему более дистальные отделы тонкой кишки и пригодному для осуществления кишечного пищеварения. Именно в двенадцатиперстной кишке и начинается процесс кишечного пищеварения. Другая важнейшая функция двенадцатиперстной кишки состоит в инициации и регулировании секреции панкреатических ферментов и жёлчи в зависимости от кислотности и химического состава поступающей в неё пищевой кашицы. Третья важнейшая функция двенадцатиперстной кишки заключается в поддержании обратной связи с желудком — осуществлении рефлекторного открывания и закрывания привратника желудка в зависимости от кислотности и химизма поступающей пищевой кашицы, а также регулировании кислотности и пептической активности секретируемого в желудке сока через секрецию гуморальных факторов, влияющих на секреторную функцию желудка.
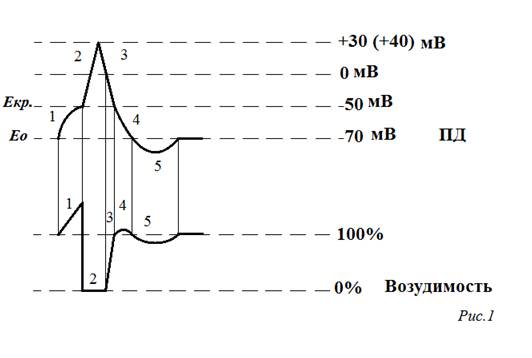
3. Современные представления о механизмах периодичности дыхания (генератора центрального инспираторного возбуждения, механизм выключения инспирации, пневмотаксический центр, механорецепторы легких) Главной особенностью дыхательного центра является периодичность, при которой возбуждение нейронов изменяется их торможением. В основе периодичности лежит функция бульбарного отдела. При этом решающая роль принадлежит нейронам дорсального ядра. Считают, что они являются своеобразным «водителем ритма». С бульбарного центра поступает возбуждение от многих образований ЦНС, в том числе от пневмотаксичного центра. Так, если перерезать ствол мозга, отделив варолиев мост от продолговатого мозга, то у животных снижается частота дыхательных движений.. Пневмотаксичний и бульбарный центры имеют двусторонние связи, с помощью которых пневмотаксичний центр ускоряет возникновение следующих инспираций и експираций. На активность нейронов дыхательного центра влияют другие отделы ЦНС, такие, как ретикулярная формация, гипоталамус, кора больших полушарий. Например, характер дыхания меняется при эмоциях. Скелетные мышцы, которые участвуют в дыхании, часто выполняют и другие движения. Да и собственное дыхание, его глубину и частоту человек может менять сознательно, что свидетельствует о влиянии на дыхательный центр коры больших полушарий. Благодаря этим связям дыхания сочетается с выполнением рабочих движений, речевой функцией человека. Таким образом инспираторных нейронов, как «водители ритма», существенно отличаются от настоящих пейсмекерных клеток. При возникновении ритмики основных дыхательных нейронов дорсального ядра необходимо учитывать два условия: а) «последовательность поступления» каждой группы нейронов именно этого отдела б) обязательное импульсацию от других отделов ЦНС и импульсацию от различных рецепторов. Дыхание является вегетативной функцией, а выполняется скелетными мышцами. Поэтому механизмы его регуляции имеют общие черты с механизмами регулирования деятельности как вегетативных органов, так и скелетных мышц. Потребность в постоянном дыхании обеспечивается автоматически за счет активности дыхательного центра. Однако вследствие того, что дыхание осуществляют скелетные мышцы, возможны и произвольные изменения характера дыхания, обусловленные влиянием коры больших полушарий на дыхательный центр. Если во внутренних органах (сердце, кишки) автоматизм обусловлен лишь свойствами водителей ритма, то в дыхательном центре периодическая деятельность управляется сложными механизмами. Периодичность обусловлена: 1) согласованной активностью различных отделов дыхательного центра, 2) поступлением сюда импульсов от рецепторов, 3) поступлением сигналов от других отделов ЦНС, в том числе и от коры головного мозга. Кроме того, при анализе механизма периодичности дыхания нужно учесть, что спокойное и форсированное дыхание существенно отличаются по количеству мышц, участвующих в этом акте. Во многом эта разница определяется уровнем привлечения вентрального отдела бульбарного дыхательного центра, в котором есть как инспираторная, так и экспираторная нейроны. При спокойном дыхании эти нейроны относительно малоактивны, а при глубоком дыхании их роль резко возрастает. 4. Билет №9 1.Уровень поляризации клеточных мембран как регулятор функционального состояния ткани. Зависимость возбудимости от потенциала покоя. Изменение возбудимости при потенциале действия. Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин: 1) неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов К, снаружи его мало. Ионов Na и ионов Cl больше снаружи, чем внутри. 2) избирательной проницаемости мембраны для ионов.. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ. Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала.
2. Сосудистотромбоцитарный гемостаз. Значение. Механизмы. Роль: 1) обеспечивает остановку кровотечения из сосудов микроциркулярного русла и в сосудах с низким АД; 2) является предфазой коагулляционного гемостаза. Фазы. 1 Рефлекторный спазм поврежденных сосудов. Обеспечивается БАВ, которые выделяются из разрушенных тромбоцитов (серотонин, НА, Адр.) – временно прекращают кровотечение. Эта реакция увеличивается при охлаждении поврежденного участка. 2 процесс. Спазм сосудов дополняется: адгезией тромбоцитов. В силу электростатического взаимодействия (тромбоцит “- „), обнажаются волокна коллагена стенки «+», происходит прилипание тромбоцитов к стенке (3 – 10с). 3 стадия. Обратимая агрегация (скучивание) тромбоцитов. Начинается почти одновременно с адгезией. Катализатор этого процесса АДФ, выделяемая из поврежденных тканей сосуда – внешняя АДФ, из тромбоцитов и эритроцитов – «внутренняя». Образуется рыхлая тромбоцитарная пробка, пропускающая плазму – белый тромб. 4 стадия.Необратимая агрегация – тромбоцитарная пробка становится непроницаемой для плазмы. Происходит это под влиянием тромбина, который меняет структуру мембраны тромбоцитов, и они сливаются в гомогенную массу. 5 Ретракция белого тромба. Это сокращение и уплотнение белого тромба, за счет сокращения нитей фибрина. Этим путем (сосудисто-тромбоцитарным) останавливается кровотечение из сосудов МЦР за 3 – 4 минуты при бытовых травмах. 3. Функциональная система поодежания газового состава крови. Резервы дыхательного процесса. Дыхание при деятельности. Дата добавления: 2015-12-15 | Просмотры: 894 | Нарушение авторских прав |