|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Химическая природа гормонов и общие закономерности молекулярных механизмов действия гормонов различной химической природыВ зависимости от химической природы гормоны классифицируют на: Ø белково-пептидные гормоны (гормоны гипоталамуса, аденогипофиза, С-клеток щитовидной железы, паращитовидных желез, островков Лангерганса поджелудочной железы) Ø гормоны – производные аминокислот (тиреоидные гормоны и катехоламины (производные аминокислоты тирозина), серотонин и мелатонин эпифиза (производные аминокислоты триптофана)) Ø стероиды (половые гормоны и гормоны коркового вещества надпочечников (кортикостероиды)).
Свое действие на организм гормоны реализуют в конечном итоге на клеточном уровне через посредство определенных рецепторов. Клеточный рецептор к гормону представляет собой, как правило, белковую молекулу (либо в более сложном случае комплекс белка с углеводом (гликопротеид)). Локализация рецепторов к гормонам в клетках-мишенях различна и во многом зависит от химической природы гормона, а, следовательно, и его способности или неспособности проникать внутрь клетки. Так, стероидные и тиреоидные гормоны проникают через плазматическую мембрану внутрь клетки, оказывают свои эффекты на клеточном уровне через ее генный аппарат (изменение трансляции определенных генов) и соответственно рецепторы для них находятся в цитоплазме клетки-мишени и ее ядре. Взаимодействие комплекса гормон-рецептор с определенными участками генома клетки-мишени обуславливает изменение конформации хроматина (его деконденсацию), в результате чего определенные гены становятся доступными для считывания информации. С этих генов начинается экспрессия определенных белков (белков-ферментов, в результате чего изменяется интенсивность метаболизма в клетке, структурных белков, белков ионных каналов и ионных насосов). Таким образом, стероидные и тиреоидные гормоны, реализующие свои эффекты в основном через генетический аппарат клетки, способны оказывать специфическое влияние на экспрессию клеточного генома, а, следовательно, прежде всего на структуру клеток-мишеней (вызывать определенные морфологические и связанные с ними функциональные перестройки). Такие гормоны, реализующие свое влияние на клетку через ее геном, оказывают морфогенетическое действие на организм. Вместе с тем эти гормоны могут действовать на клетку-мишень и негеномным путем (непосредственно оказывая влияние на определенные внутриклеточные структуры, активность ферментов и состояние ионных каналов и насосов). Так, тиреоидные гормоны, наряду с влиянием на экспрессию генома в клетках-мишенях, способны оказывать и непосредственное (негеномным путем) регулирующее действие на те или иные клеточные структуры (в частности, активность Са2+-АТФазы саркоплазматического ретикулюма в скелетных и сердечных мышечных волокнах, активность аденилатциклазы и уровень цАМФ в мышечных волокнах, тимоцитах и эритроцитах, продолжительность нахождения потенциалзависимых Na+-каналов в открытом состоянии в момент деполяризации мембраны возбудимой структуры и т.д.).
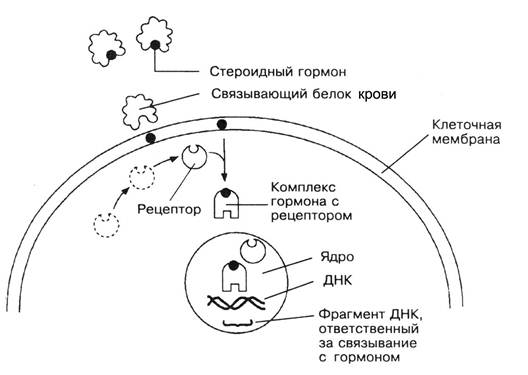
Рис. Взаимодействие стероидного гормона с клеткой. Транспорт стероидных гормонов во внутренней среде осуществляют специальные белки. Стероидный гормон отделяется от связывающего белка и проходит через клеточную мембрану внутрь клетки, где соединяется с рецептором. Комплекс гормона с рецептором поступает в ядро и взаимодействует со строго определенным фрагментом ДНК с последующей активацией конкретных генов
Катехоламины и большинство белково-пептидных гормонов не способны проникать внутрь клетки, в связи с чем реализуют свои эффекты на клеточном уровне через рецепторы встроенные в плазматическую мембрану и специальные внутриклеточные посредники (цАМФ, цГМФ, ионы Са2+, инозитолтрифосфат, диацилглицерол и т.д.). Действие большинства белково-пептидных гормонов и катехоламинов на клетку сопряжено с активацией определенных мембранносвязанных ферментов, что приводит к резкому повышению уровня соответствующих внутриклеточных посредников (вторичных мессенджеров действия гормона на клетку). Так, для катехоламинов и некоторых белково-пептидных гормонов (глюкагон, рилизинг-факторы гипоталамуса и некоторые другие) характерен аденилатциклазный механизм действия (раскрыт Сазерлендом и Робинзоном), сущность которого состоит в следующем: 1. гормон связывается с рецептором, встроенным в плазматическую мембрану 2. образовавшийся комплекс "гормон-рецептор" активирует расположенный по соседству в мембране мембранносвязаный фермент аденилатциклазу. Причем информация от комплекса "гормон-рецептор" на аденилатциклазу передается через особый регуляторный G-белок, встроенный в мембрану (этот белок активируется путем присоединения ГТФ) 3. активная аденилатциклаза катализирует превращение АТФ в 3',5'-циклическую АМФ (цАМФ), что приводит к резкому повышению уровня цАМФ в цитоплазме клетки-мишени (одна молекула связанного с рецептором гормона приводит к образованию около 500 молекул цАМФ) 4. цАМФ активирует фермент протеинкиназу путем отсоединения от нее двух ингибиторных субъединиц 5. активная протеинкиназа путем ограниченного протеолиза активирует ряд других ферментов (липазу, фосфорилазу и т.д.), что сопровождается изменением интенсивности метаболизма клетки-мишени, а, следовательно, отражается и на ее функциональном состоянии. Кроме того, цАМФ может влиять на состояние определенных ионных каналов в мембранах клетки-мишени (плазматической, саркоплазматического ретикулюма, митохондриальной), изменяя их проницаемость для тех или иных ионов 6.
Рис. Схема аденилатциклазного механизма действия гормонов
Внутриклеточными посредниками в действии некоторых белково-пептидных гормонов могут выступать инозитол-1,4,5-трифосфат и диацилглицерол (инозитолфосфатный механизм действия гормонов). Сущность этого механизма во многом сходна с таковой для аденилатциклазного механизма (его результатом также является активация протеинкиназы, активирующей ряд других ферментов), а отличия касаются природы вторичных посредников действия гормона: 1. гормон связывается с рецептором, встроенным в плазматическую мембрану 2. комплекс "гормон-рецептор" активирует G-белок путем присоединения к нему ГТФ 3. активный G-белок активирует фосфолипазу С, которая катализирует гидролиз фосфатидилинозитолполифосфата, в результате чего высвобождаются инозитолтрифосфат и диацилглицерол (внутриклеточные посредники действия гормонов) 4. водорастворимый инозитол-1,4,5-трифосфат поступает в цитозоль и вызывает высвобождение ионов Са2+ из внутриклеточных депо, а диацилглицерол диффундирует в липидной фазе плазматической мембраны к расположенной на ее внутренней поверхности Са2+-зависимой протеинкиназе С и активируя ее с участием фосфатидилсерина в качестве кофактора (диацилглицерол путем индукции конформационных перестроек в фосфолипазе С повышает ее чувствительность к кальцию, а активируется данный фермент самим свободным кальцием, концентрация которого в цитоплазме клетки-мишени повышается под действием инозитолтрифосфта) 5. ионизированный кальций, концентрация которого увеличивается в цитоплазме клетки под инозитолтрифосфата, выступает в роли посредника и способен стимулировать мышечное сокращение в мышечных клетках, активировать кальций-зависимую фосфокиназу, которая путем фосфорилирования ряда внутриклеточных ферментов активирует их 6. активированная под действием диацилглицерола и кальция мембранная протеинкиназа С путем фосфорилирования активирует ряд других белков-ферментов или изменят состояние белков ионных каналов. 7. инактивация инозитолтрифосфата и диацилглицерола в клетке, завершающаяся в конечном итоге ресинтезом фосфатидилинозитолполифосфата, осуществляется путем определенных метаболических реакций, происходящих с участием клеточных ферментных систем. В частности, инозитолтрифосфат последовательно дефосфорилируется до инозитола, а диацилглицерол конвертируется в фосфатидную кислоту, после чего – в цитозиндифосфат-диацилглицерол, который соединяется с инозитолом (продуктом дефосфорилирования инозитолтрифосфата), образуя фосфатидилинозитол и завершая тем самым рабочий цикл.
Рис. Схема инозитолфосфатного механизма действия гормонов
Системы вторичных посредников – цАМФ и инозитолтрифосфата – являются высокоэффективными биологическими усилителями, преобразующими реакцию между гормоном (или медиатором) и мембранным рецептором в фосфорилирование множества внутриклеточных белков (которые затем могут влиять на течение метаболических процессов и функциональные отправления клетки), а также временное повышение концентрации свободного ионизированного кальция в цитоплазме, также способного вызывать определенные метаболические и функциональные перестройки в клетке. Причем эти две тесно связанные регуляторные системы используются многими внеклеточными веществами (гормонами, медиаторами, нейропептидами и т.д.) для регуляции разнообразных внутриклеточных процессов. Так, например, путем активации инозитолфосфатного механизма действуют ацетилхолин, серотонин, АДГ и тиреотропный гормон, а впервые в онтогенезе организма рецептор системы инозитолфосфата активируется в мембране яйцеклетки спермием, и инозитолтрифосфат принимает участие в регуляторных реакциях, сопровождающих оплодотворение яйцеклетки. Аденилатциклазная и инозитолфосфатная внутриклеточные мессенджерные системы тесно взаимодействуют друг с другом, что позволяет им осуществлять тонкую регуляцию клеточных функций. Таким образом, большинство белково-пептидных гормонов и катехоламинов способны влиять на активность ряда ключевых ферментов клетки-мишени, а, следовательно, изменять интенсивность и направленность обмена веществ в ней (т.е. вызывать метаболические и функциональные перестройки). Вместе с тем инсулин и гормон роста, несмотря на то, что оказывает свое действие на клетки-мишени через посредство рецепторов, встроенных в плазматическую мембрану, способны через посредство внутриклеточных посредников активировать в клетках-мишенях общий белковый синтез (т.е. в отличие от стероидных и тиреоидных гормонов, дерепрессирующих строго определенные гены и в результате этого стимулирующих синтез строго определенных белков в клетке, инсулин и гормон роста неспецифически усиливают синтез большинства синтезируемых в клетке белков путем повышения проницаемости мембран клетки-мишени для аминокислот или активности ферментов, участвующих в процессе транскрипции и трансляции). Продолжительность латентного (скрытого) периода действия гормона во многом зависит от механизмов его действия на клеточном уровне. В частности, эффекты белково-пептидных гормонов и катехоламинов, не связанные с активацией экспрессии генома и реализующиеся через посредство мембранных рецепторов и специальных внутриклеточных посредников, начинают проявляться спустя очень короткий латентный период (при внутривенном введении гормона – через 5-10 минут). Эффекты же тиреоидных и стероидных гормонов, предполагающие активацию клеточного генома, начинают проявляться спустя гораздо более длительный латентный период (24-48 часов и более). Гормоны, поступившие в периферическую кровь, могут находиться в ней в двух состояниях: Ø химически связанном виде (преимущественно с белками плазмы крови). Большая часть молекул гормона в крови, как правило, химически связана с определенными белками плазмы и представляет собой своеобразный резерв гормона, который может быть использован организмом в случае резкого повышения потребности в нем. Сами же белки плазмы крови, связывающие и транспортирующие гормоны, представляют собой как бы своеобразный буфер для них Ø свободной форме (именно химически несвязанные молекулы гормона способны проникать в ткани и, взаимодействуя с определенными рецепторами, оказывать влияние на клетки-мишени). Причем между химически связанной и свободной формами гормона существует состояние динамического равновесия. Так, уменьшение концентрации свободной формы гормона в крови (возникающее, например, в результате усиленного потребления его тканями) приводит к усиленному высвобождению его из химически связанного состояния и, как следствие, нормализации концентрации свободной формы гормона в циркулирующей крови. Дата добавления: 2015-02-06 | Просмотры: 4449 | Нарушение авторских прав |
 наконец, концентрация цАМФ в клетке контролируется фосфодиэстеразой, катализирующей распад цАМФ до неактивной АМФ, что обеспечивает дискретность действия гормона на клетку. В свою очередь, активность фосфодиэстеразы определяется комплексом ионов Са2+ с кальмодулином.
наконец, концентрация цАМФ в клетке контролируется фосфодиэстеразой, катализирующей распад цАМФ до неактивной АМФ, что обеспечивает дискретность действия гормона на клетку. В свою очередь, активность фосфодиэстеразы определяется комплексом ионов Са2+ с кальмодулином.
