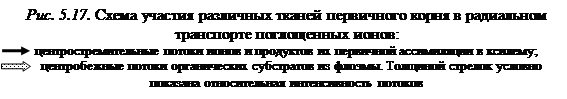|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Транспорте веществ
При радиальном движении минеральных веществ из окружающей среды к сосудам ксилемы они проходят через соответствующие ткани корневой системы. Первой такой тканью является ризодермис. Ризодермис (эпидермис) – поверхностная ткань клеток корней. В этом случае нужно ответить на вопрос: поступают ли ионы в клетки ризодермиса непосредственно с внешнего окружения, а затем в клетки коры, или поступают в кору из межклеточного пространства. После многочисленных опытов и дискуссий был сделан вывод, что функционально первой тканью поступления ионов является ризодермис, а клетки корней, которые омываются растворами свободного пространства, не играют значительной роли в обеспечении растений питательными веществами. Чтобы подтвердить этот вывод приведем только один пример. В контрольных опытах (когда отсеченные корни поглощали Rb+ из раствора) в корнях ячменя меченый рубидий находили во всех клетках коры, вплоть до самой эндодермы. В опытах с корнями, в которых симпластические связи были разрушены плазмолизом, Rb+ приблизительно в тех же количествах накапливался в периферийных клетках, но практически не поступал в кору. Таким образом, в обычных условиях, при относительно низких концентрациях ионов в окружающей среде, их первичное поступление происходит, главным образом, в ризодермис – с этим сейчас согласны все. Спор идет только по вопросу обусловлено ли это тем, что корневой ризодермис имеет позиционное или функциональное преимущество? Д. Б. Вахмистров с сотрудниками считает, что корневой ризодермис владеет более развитой и активной системой мембранных транспортных механизмов. Возникает еще вопрос: происходит поступление во все ризодермальные клетки, или происходит поступление локально, главным образом в корневые волоски. Наиболее распространенным является мнение, что мембраны корневых волосков и безволосковых клеток функционально одинаковы; корневые волоски усиливают поступление, поскольку они увеличивают общую поглощающую поверхность. Ряд исследователей считает, что это заключение не полно отражает ситуацию; было сделано уточнение. На основе новых экспериментальных результатов было сделано уточнение. Оно заключалось в том, что вероятно, корневой волосок является, с одной стороны, основным «входом» для поступающих ионов в симпласт, а с другой – основным «коридором», по которому эти ионы проходят по симпластичному пути в кору и дальше к сосудам ксилемы. Но с возрастом корневые волоски теряют это преимущество. Загрузка симпласта происходит, таким образом, в ризодермисе при участии Н+-помпы. Следующая за ризодермисом ткань – кора, которая, как мы отметили, не принимает значительного участия в поступлении ионов. Тогда какую же роль она выполняет? На пути от наружной среды до сосудов ксилемы – это единственная многоклеточная ткань корня. Почему? Вероятно, кора существует в корнях для того, чтобы разделить ризодермис и сосуды ксилемы и направить ионы по достаточно длинному пути, чтобы дать им время быть ассимилированными. С этой точки зрения, вероятно, что она главный метаболический реактор. Подтверждением такого вывода могут служить данные, полученные в опытах по выращиванию томатов при разных условиях аэрации: в воздушной, а также водных культурах при непрерывном продувании и без продувания (рис. 5.15).
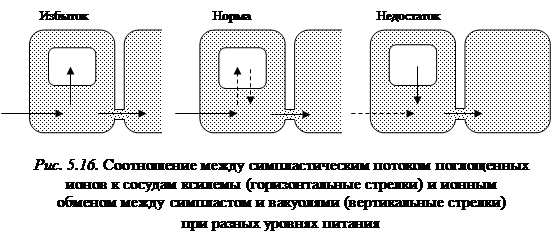
Оказалось, что чем хуже аэрация, тем больше площадь коры и соответственно меньше площадь центрального центра. По мере ухудшения аэрации отношение кора/стела значительно повышалось. Вместе с уменьшением центрального цилиндра (стела) снижалось суммарное сечение сосудов. Эти результаты можно интерпретировать следующим образом. При хорошей аэрации корней (аэропоника) метаболизм протекает интенсивно и для усвоения поступающих ионов не требуется длительного времени, а следовательно, большой протяженности пути до сосудов. По мере ухудшения аэрации метаболизм становится менее интенсивным и для ассимиляции ионов требуется все большее время и более длительный путь, т. е. необходимо увеличение толщины коры корня. Кора выполняет и другую функцию – функцию транспортного «буфера», регулирующего поток ионов по радиусу корня. На пути этого потока происходит конкуренция между двумя транспортными потоками: магистральным симпластическим путем к сосудам и боковым тупиковым путем в вакуоль. При излишке питания – избыток из симпластического потока направляется в вакуоль, – к сосудам подается нормальное количество питательных веществ (рис. 5.16).
В норме ионные потоки на тонопласте уравновешены и практически все поступающие ионы направляются по симпласту прямо в сосуды. При недостатке равновесие сдвигается в сторону симпласта, дефицит пополняется за счет ионов, ранее запасенных в вакуолях, – и к сосудам вновь подается нормальное количество минеральных веществ. Благодаря пояскам Каспари эндодерма выполняет в корнях две функции. С одной стороны, она служит диффузионным барьером, который не пропускает апопластические ионы из коры в центральный цилиндр; с другой, она работает как осмотический барьер, локализуя корневое давление и ксилемный сок внутри центрального цилиндра, и не дает ксилемному соку вытекать, выдавливаться под воздействием корневого давления в окружающую среду и бесполезно растрачиваться для растения. Необходимо обратить внимание на следующий факт. Являясь диффузионным и осмотическим барьером, эндодерма делит апопласт корней на две части: кортикальный и стелярный. Первый контактирует с наружной, как правило низкосолевой средой (концентрация, например, калия ~ 1 мМ). Стелярный же апопласт соединяется с ксилемным соком и омывается высокосолевым раствором (концентрация калия ~ 50 мМ). Это значит, что клетки снаружи эндодермы и внутри находятся в разном ионном окружении. Таким образом, кора и ризодермис представляют собой как бы пресный чехол, который служит предохранительной прокладкой между низкосолевым почвенным раствором и всеми остальными клетками растений, которые функционируют в высокосолевом растворе.
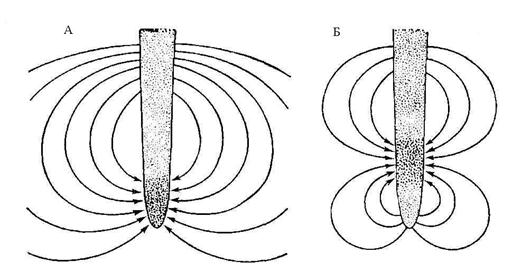
За эндодермой идет однослойная ткань – перицикл. До последнего времени ее рассматривали как мерестематический очаг заложения боковых корней. Между тем она является как бы тропинкой, ведущей к сосудам. Сосуды протоксилемы или тесно примыкают к кольцу клеток перецикла (например, у кукурузы), либо непосредственно включены в это кольцо (например, ячмень). На основе анализа размещения и количества плазмодесм между клетками перецикла и соседними тканями был сделан вывод, что перецикл работает как кольцевой коллектор. Он подбирает ионы, которые идут из коры через эндодерму, и направляет их по короткому пути (по дуге) к сосудам ксилемы. Разгрузка питательных веществ в сосуды ксилемы происходит пассивно, без участия помпы; помпа участвует только в обратной реабсорбции ионов ксилемного сока, стабилизируя его состав. Общая схема участия тканей корня в радиальном транспорте ионов представлена на рис. 5.17. Размещение транспортных систем в плазматической мембране клеток корня. Долгое время плазматическую мембрану растительной клетки рассматривали как изотропную поверхность, на которой транспортные механизмы размещены равномерно. Но в последнее время появились основания полагать, что эта поверхность анизотропна, что транспортные системы размещены неравномерно, при этом эта неравномерность определенным образом упорядочена, т. е. транспортные механизмы разнесены на мембранной поверхности – сегрегированы. Еще в 1960 г. Скот с сотрудниками сообщили, что растущие корни (конские бобы) окружены силовыми линиями, вдоль которых во внешней среде текут сверхслабые электрические токи (рис. 5.18).
Долго эти результаты не удавалось повторить. Но в 70–80 гг. ХХ века в одной из лабораторий Калифорнии был разработан специальный вибрирующий электрод. С его помощью появилась возможность регистрировать эти малые по силе трансклеточные электрические токи. Трансклеточные электрические токи сейчас выявлены, у множества самых разнообразных организмов – как одно-, так и многоклеточных. Основным носителем выходящего тока оказался протон. Ток, который входил в клетку, осуществлялся ионами K+. Во всех растительных клетках токи всегда входили в растущую часть клетки и выходили из противоположной – не растущей. По теперешним представлениям, как мы уже подчеркивали, всю многочисленность мембранных механизмов можно свести к двум основным категориям: активный транспорт (помпы) и пассивный транспорт (ионные каналы и переносчики). Каналы катионной проводимости сконцентрированы в растущем конце корневых волосков, а анионные – в не растущем основании. Как мы уже говорили, ток положительных зарядов, выходящих из клетки, это протонный ток. Таким образом, насосы, вероятно, размещены у основания. Более того, было показано, что РЭП на мембране основного корня больше, чем на мембране выроста корневого волоска. Поэтому насосы, генерирующие эту РЭП, работают где-то в теле корня. Характеристики каналов и насосов мы уже рассмотрели в разделах 5.3. и 5.4. А как расположены на поверхности анионные каналы? Были проделаны опыты с катионными и анионными красителями: анионные красители поступали в основание волосковых клеток, а не в их выросты. Что касается безволосковых клеток, то в них не поступали ни катионные, ни анионные красители. Наиболее простое и наиболее вероятное объяснение эффекта возникновения наблюдаемых катионных клеточных токов заключается в пространственном разделении (сегрегации) катионных насосов и катионных каналов в мембране. Общая картина представляется следующей (рис. 5.19). Протоны выкачиваются из клетки электрогенными насосами, размещенными в ее нерастущем основании, а ионы K+ входят за счет образовавшегося электрохимического градиента в клетку через катионные каналы, расположенные в другом месте – в растущем конце корневого волоска. Поэтому между основанием и кончиком во внешней среде возникает электрический ток. Анионные системы расположены там же, где насосы – у основания. Если бы две системы катионная и анионная находились на одинаковом расстоянии от насоса, то протон имел бы одинаковые шансы вернуться в клетку и через анионный симпортер и через катионный канал. В первом случае вход был бы полезным, поскольку он сопряжен с поступлением аниона. Во втором, он был бы бесполезным, и даже опасным, поскольку протон конкурировал бы за вход с другими питательными катионами. Клетке полезно направить протон через анионный симпортер. Поэтому, вероятно, она и удаляет катионные каналы от насоса, а анионные оставляет рядом.
Все это относится к корням, имеющим корневые волоски. В безволосковых корнях (водные культуры, ряд растений, не имеющих вообще корневых волосков – ряска, нарцисс и др.) сегрегация катионной и анионной проводимостей происходит не между частями одной клетки, а между клетками ризо- и гиподермиса (первый слой коры), которые, вероятно отличаются по своим ростовым процессам. Общий принцип сегрегации ионтранспортных систем, вероятно, универсален. Но конкретные формы проявления этого принципа в разных условиях и разных объектах могут быть весьма разнообразными. Метаболизм в корнях. Корневая система не только обеспечивает поступление воды и минеральных элементов в растение, но в ней самой происходят соответствующие метаболические реакции. Обмен веществ в корнях связан с его ролью для целого растения. Корень – это специализированный орган поступления воды и минеральных элементов из почвы. Поэтому часть процессов биосинтеза направлена, во-первых, на построение аппарата поступления (поглощения) и систем переноса, которые необходимы для транспорта в корень ионов, органических соединений и воды и доставки к местам потребления. Во-вторых, в корнях происходит полная или частичная переработка поступивших ионов и перевод их в транспортную форму (восстановление, включение в разные органические вещества). И, в-третьих, в корнях синтезируются физиологически активные вещества: фитогормоны цитокининовой природы и гиббереллины, необходимые для развития и роста растений. Особенностью метаболизма корневой системы является то, что источником углерода служат продукты фотосинтеза, которые поступают из надземных органов, главным образом, из закончивших рост листьев. Основная транспортная форма углеводов – сахароза. Сахароза обладает преимуществами по сравнению с другими соединениями своей универсальностью в процессах обмена веществ. В первую очередь это обусловлено ее метаболической инертностью, что важно при ее движении по сосудам. Метаболизация сахарозы происходит с помощью фермента инвертазы, который отсутствует в проводящих элементах и проявляет активность в других тканях. При метаболическом превращении сахарозы под действием инвертазы образуются соединения (глюкоза, фруктоза), которые используются на поддержание роста и функциональной активности корня; часть соединений входит в состав корневых выделений, а в составе пасоки поступает в надземные органы. В метаболизме клеток корня используются также поглощенные из окружающей среды минеральные вещества, вода, а также некоторые органические соединения, выделяемые микроорганизмами ризосферы: витамины, аминокислоты, хотя их роль в этих процессах, возможно, и невелика. Растения способны усваивать и другие виды моносахаров, например галактозу. Некоторые сахара участвуют в синтезе полимеров клеточных стенок (целлюлоза, гемицеллюлоза, пектиновые вещества), а также используются для синтеза крахмала в значительных количествах, откладывающихся в тканях корня. Другая часть моносахаров тратится на процессы дыхания, поставляющего энергию в форме АТФ, а также на органические кислоты, образующиеся в цикле Кребса. Их присутствие необходимо для транспорта поглощенных корнем катионов, поскольку анионы органических кислот выполняют функцию противоионов, нейтрализуя положительные заряды катионов. Другая функция органических кислот – регуляция внутриклеточного рН, которая в результате преимущественного поступления в клетку катионов щелочно-земельных металлов могла бы возрастать до пределов, нарушающих нормальную жизнедеятельность клетки. Анионы органических кислот вместе с катионами могут поступать в вакуоль, тем самым, снижая концентрацию катионов в цитоплазме и обуславливая новое поглощение их клеткой. Запасы органических кислот в клетках пополняются также в результате процессов темновой фиксации СО2, что может составить 1–5 % общего содержания углерода в растении. Значительная часть органических кислот используется в качестве акцепторов для первичной ассимиляции аммиака с образованием аминокислот. Аминокислоты и амиды служат основными транспортными формами азота, поступающими в надземные органы растения, а, кроме того, являются исходным материалом для синтеза в корне всех азотосодержащих органических соединений (белков, пуриновых и пиримидиновых оснований, нуклеиновых кислот и др.). Синтез аминокислот локализован в определенных участках корня. Максимальное количество свободных аминокислот содержится в зоне меристемы, очевидно, из-за процессов белкового синтеза, идущих в этой части корня. Интенсивность образования аминокислот меняется с возрастом растения, достигая максимума в фазе цветения. После цветения, когда количество аминокислот в пасоке снижается, содержание амидов меняется медленнее, и удельный вес последних к концу вегетации может возрасти. Существует и суточный ритм накопления в корнях и подачи аминокислот с пасокой в надземные органы. В дневные часы интенсивность синтетических процессов выше. В ночные часы в пасоке накапливаются в основном другие формы небелкового азота (аммиак, нитраты, амиды, азотистые основания), что, по-видимому, связано с изменением функционального состояния корневой системы. В корнях синтезируются содержащие азот порфирины, некоторые витамины (В1, В2, никотиновая, аскорбиновая кислоты), ростовые вещества (цитокинин, АБК, гиббереллины, алкалоиды и др.). Часть их непосредственно вовлекается в метаболические процессы корня и всего растения, а также в обменные процессы, связанные с поглощением веществ из почвы и образованием новых структур корневой системы, другие могут выделятся в почву, формируя среду ризосферы и воздействуя на микроорганизмы почвы, а также на другие виды растений. Таким образом, в корне происходят интенсивные метаболические реакции, направленные на поддержание целостности растительного организма.
Дата добавления: 2014-12-11 | Просмотры: 1306 | Нарушение авторских прав |