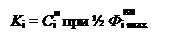|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Системы пассивного транспорта
Ионные каналы. Для прохождения ионов пассивным путем через мембраны имеются специальные структуры, которые носят название ионных каналов. Ионные каналы обеспечивают быстрый векторный перенос ионов по градиенту электрохимического потенциала. Важной особенностью транспорта через каналы является снижение энергетических затрат на трансмембранный перенос ионов. Например, для перемещения 1 моля ионов калия через липидный бислой растрачивается энергия ~250 кДж, а при наличии каналов в этом слое (калиевых) ~20 кДж. Что собой представляет ионный канал? Ионные каналы формируются интегральными (обычно субъединичными) белками. Они пронизывают мембрану таким образом, что образуется гидрофильная пора. Канал – это белковая макромолекула, образующая пору через двухслойную липидную мембрану. В общем виде схематически строение ионного канала можно представить следующим образом. Основные структурные элементы канала: устье – расширенная часть поры, которая находится по обеим сторонам мембраны; узкая часть поры – туннель (рис. 5.3).
В стенах туннеля с внутренней стороны размещены атомы кислорода. Считают, что конформация канала меняется в ответ на физиологические раздражители, которыми могут быть электрическое поле, химические соединения (медиаторы и др.). Эти конформационные изменения обычно называют воротными. Воротными их называют потому, что, открывая и закрывая канал, они обуславливают движение проникающих через мембрану ионов. Опыты показали, что ворота расположены ближе к внутренней стороне мембраны. Механизмы, которые управляют воротами, т. е. открытием и закрытием канала, получили название сенсоров. Сенсоры могут действовать под влиянием либо электрического поля на мембране, либо химических агентов (медиаторов). Одной из важнейших характеристик ионных каналов является селективность, обусловленная особенностью их строения: диаметром канала, природой и размещением заряженных групп. Большое значение при этом имеют заряды, локализованные непосредственно у входа в канал, где они выполняют функцию селективного фильтра. Открытая пора дает возможность ионам проходить через мембрану в водном окружении. При этом ион преодолевает ряд энергетических барьеров, задерживаясь в местах связывания на различные промежутки времени. Подобно ферментам, каналы выполняют роль катализаторов, селективно увеличивая скорость движения ионов на много порядков в сравнении с движением без катализаторов. Наибольшим лимитирующим энергетическим барьером является «селективный фильтр». Этот фильтр такой узкий, что ионы должны хоть бы частично дегидратироваться. Каким образом происходит дегидратирование ионов в канале? Мы подчеркивали, что внутренняя часть канала содержит атомы кислорода. От характера взаимодействия гидратной оболочки иона со стенками поры, содержащими атомы О2, зависит перенос иона через мембрану. Ионы, величина которых больше диаметра поры не проходят по стерическим эффектам. Ионы, величина которых значительно меньше диаметра поры не проходят (или плохо проходят) из-за того, что недостаточно эффективно взаимодействуют с атомами кислорода. Когда ион связывается не со всеми атомами О2 (О2 не может подойти из-за упругости стенок), то его потенциальная энергия выше, чем в воде и ион не может пройти через канал. И только тот ион, который эффективно взаимодействует с атомами О2 (последний как бы занимает место воды вокруг иона), хорошо проходит через пору соответствующего диаметра (рис. 5.4).
Таким образом, в канале имеется узкий туннель с селективным фильтром вблизи наружной стороны мембраны, воротное устройство вблизи внутренней стороны. Сенсор, расположенный в липидном бислое, управляет воротами под действием внутримембранного поля или хемомедиаторов. Более широкая часть канала – устье расположено с двух сторон мембраны и способствует более большим по своему диаметру ионам блокировать прохождение основных ионов. На мембране также существует небольшой поверхностный заряд (отрицательный), который обуславливает образование так называемого двойного (неперемешивающего). Этот слой может быть очень значительным по своим размерам и влиять на движение ионов через мембрану (одна из причин, почему неизвестна величина Δ х). В настоящее время на мембранах растительных клеток идентифицированы несколько видов ионных каналов. В частности это Са2+-Na+ канал и K+-каналы. Са2+-Na+ канал идентифицирован в нестационарных условиях, т. е. при возбуждении клетки и возникновении ПД. Повышенный интерес к Са2+-каналам связан с особой ролью Са2+ как вторичного посредника во многих функционально важных процессах. Са2+-Na+-канал плазмалеммы является потенциалзависимым и для его функционирования необходимо чтобы внутри клетки находился Mg2+ и АТФ; существует также цАМФ-зависимая регуляция функционального состояния Са2+-Na+-каналов растительной клетки. Что касается калиевых каналов, то они изучены более детально. Выявлен ряд их свойств: механизм движения ионов в канале, потенциалзависимость, блокирование и т. д. Установлено, что калиевые каналы бывают двух типов: Д-каналы и Г-каналы. Д-каналы в обычных условиях активируются в интервале потенциалов от –80 до –100 мв, а Г-каналы – от –100 мв и ниже. Ряд проницаемости качественно одинаков для обоих видов K-каналов: K+ > Rb+ > NH4+ > Na+ > Li+ > Cs+
K+ > Mg2+ > Sr2+ > Ca2+ > Cd2+ > Ba2+ Известно достаточное количество небольших гидрофобных молекул, повышающих проницаемость мембран к ионам (ионофоры). Существует два вида ионофоров – каналообразующие ионофоры (каналоформеры) и подвижные переносчики. Примером каналообразующего ионофора является грамицидин А – линейный пептид, состоящий из 15 аминокислот. Вероятно, в липидном бислое две молекулы грамицидина объединяются и образуют трансмембранный канал, позволяющий моновалентным катионам передвигаться по электрохимическим градиентам. Кроме катионных каналов на плазматических мембранах растительных клеток идентифицированы и анионные каналы. В плазмалемме и тонопласте харовых водорослей и клетках высших растений присутствуют анионные каналы, активируемые сдвигом потенциала в сторону гиперполяризации и повышением уровня Са2+. Есть и анионные каналы, активирующиеся при деполяризации мембраны. В замыкающих клетках устьиц эти каналы открываются, например, при мембранном потенциале от –80 до –20 мв и являются также Са2+-зависимыми. У высших растений на тонопласте обнаружены медленные и быстрые вакуолярные каналы. Для анионных каналов первого типа характерна высокая проницаемость к катионам, что объясняется совместным передвижением аниона с катионам, поскольку селективный фильтр канала, как считают, представлен анионной группой. Второй тип анионных каналов более специфичен по отношению к анионам по сравнению с катионами и лучше проницаем для нитрата, чем для хлорида. На открывание и закрывание ионных каналов влияют изменения мембранного потенциала, ионы Са2+, механические воздействия, фосфорилирование и дефосфорилирование белков каналов. Система облегченного транспорта (переносчики). Возможность принимать участие молекул переносчиков в процессе транспорта веществ через мембраны растительных клеток было высказано впервые Остерхаутом в 30-х гг. ХХ в. В настоящее время концепция переносчиков довольно широко используется для интерпретации экспериментальных результатов.
Процесс передвижения переносчика с ионом и самого переносчика с одной стороны мембраны на другой, который проходит без затраты энергии по законам диффузии, называют облегченной диффузией. В основе облегченной диффузии лежит обратное связывание транспортируемого иона с белком переносчиком, который проходит через мембраны с освобождением иона на другой ее стороне. Переносчики бывают подвижные и фиксированные (рис. 5.5). Облегченная и простая диффузия отличаются одна от другой, по меньшей мере по трем критериям: 1. Ионный поток при простой диффузии, как правило, пропорционален концентрации иона с той стороны мембраны, откуда начинается его движение. При облегченной диффузии мы видим отклонение от линейности: зависимость скорости транспорта (потока) от концентрации имеет выход на плато, т. е. происходит насыщение (рис. 5.6). Это связано с тем, что количество переносчиков в мембране ограничено, поэтому в какой-то момент все переносчики оказываются заполненными переносимыми ионами. 2. Переносчики, функционирующие по механизму, облегченной диффузии, владеют, как правило, выраженной специфичностью – селективно транспортируют одни ионы и не взаимодействуют с другими. 3. Облегченная диффузия чувствительна к ингибиторам – веществам, способным избирательно инактивировать переносчик. Процесс поступления ионов с помощью переносчиков описывается в рамках клетки Михаэлиса – Ментен (см. рис. 5.6).
где K i – константа Михаэлиса;
Следует отметить, что константа K i характеризует сродство переносчика к переносимому иону. Чем ниже K i, тем больше сродство, т. е. ион связывается с молекулой переносчика. В качестве примера облегченной диффузии часто приводят функционирование высокоселективного ионофора валиномицина – подвижного переносчика K+. Валиномицин – антибиотик депсипептидной природы. Молекула валиномицина построена из трех идентичных фрагментов, в каждый из которых входит остаток D-валина, L-лактата, L-валина и D-оксиизовалериановой кислоты. Эти остатки формируют в центре молекулы отрицательно заряженную гидрофильную полость. Наружная часть молекулы гидрофобна. Стерически внутренняя часть молекулы валиномицина точно соответствует дегидратированному иону K+. При образовании комплекса K+-валиномицин происходит нейтрализация внутримолекулярных зарядов ионофора положительным зарядом K+, комплекс становится более гидрофобным и поэтому легко проходит через мембрану, транспортируя ион K+.
Дата добавления: 2014-12-11 | Просмотры: 1794 | Нарушение авторских прав |




 Скорость поступления () иона в клетку пропорциональна наружной концентрации этого иона в интервале низких концентраций, а затем снижается по мере роста и в конце концов приближается к максимуму (насыщение). В этом случае поток записывается в виде:
Скорость поступления () иона в клетку пропорциональна наружной концентрации этого иона в интервале низких концентраций, а затем снижается по мере роста и в конце концов приближается к максимуму (насыщение). В этом случае поток записывается в виде: