|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
И флоэмного сока у однолетнего люпина
Органические компоненты изменяются в зависимости от вида растения и от природы ионов, которые присутствуют в почвенном растворе. Если поместить корни в солевой раствор, из которого катионы поступают лучше, чем анионы (например, в раствор сернокислого калия), то количество поступивших катионов в корнях будет выше количества анионов. Растения компенсируют эту несбалансированность синтезом органических анионов, обычно карбоновых кислот. При своем движении по сосудам ксилемы минеральные вещества взаимодействуют с клеточными оболочками. В клеточных оболочках, как мы знаем, имеются многочисленные участки с фиксированными отрицательными зарядами, способными связывать катионы, в частности Са2+. Когда в листе ионы кальция переходят из ксилемы в мезофилл, в клеточных стенках ксилемы некоторые участки освобождаются. Освободившиеся участки через некоторое время занимаются кальцием из участков, размещенных ниже, или Са2+ из ксилемного сока. Поэтому считают, что перемещение Са2+ по ксилеме напоминает восходящее движение по ионообменным участкам. Кальций слабоподвижный элемент. Его подвижность увеличивается, когда он поступает в виде хелатного или еще какого-либо комплекса. Нужно отметить, что часть ионов, которая попала в ксилему, не достигает стебля и листьев. Эти ионы выходят из ксилемного сока по пути их движения в побег и листья. Выход ионов наблюдается и в тканях корня, и в тканях стебля и др. (рис. 5.20).
Если использовать гуттирующие растения, можно определить какое количество солей, находящихся в ксилемном соке корней, выходит из него за время перемещения сока по побегу. Очень часто концентрация в гутируемой жидкости намного ниже, чем в пасоке, вытекающей из отделенных корней. После того, как ксилемный сок достигнет мельчайших разветвлений проводящей системы листа, вода и растворенные в ней вещества поступают как в оболочки клеток (апопласт), так и в цитоплазму клеток мезофилла. Большая часть воды испаряется. В результате испарения воды из клеток мезофилла листа возрастает концентрация солей в растворе, заполняющем поры клеточных оболочек. В конечном итоге может возникнуть небольшой градиент концентрации, достаточный для того, чтобы обусловить обратную диффузию в ксилему навстречу массовому потоку. Вообще, поступление минеральных элементов из апопласта в клетки листа происходит в результате активной работы Н+-помпы. Растущие листья являются мощными акцепторными зонами для минеральных элементов ксилемного сока. Если вспомнить, что объем воды, испаряемый за день в процессе транспирации, в 5–10 раз превышает объем клеток листа, то станет ясно, что должен существовать какой-то механизм, удаляющий излишек солей, чтобы осмотическое давление клеток не достигло опасного уровня. Существуют три таких механизма: 1. Образование тяжелорастворимых осадков солей. 2. Соль удаляется из листа с той же скоростью, как и поступает. 3. Соль накапливается в специальных клетках – солевых железках. Большинству растений, которые обитают на незасоленных почвах, присущи два первых механизма. Третий механизм присущ растениям, произрастающим на засоленных почвах. В процессе эволюции у этих растений сформировалось несколько типов солевых желез: в одних случаях соль скапливается в особых пузыревидных клетках; в других, она непрерывно выделяется на поверхность листа через железистые клетки, а затем смывается дождем или сдувается ветром. Железистые клетки должны обладать чрезвычайно прочными стенками для того, чтобы противостоять огромному гидростатическому давлению (тургору), ибо внутриклеточная концентрация солей может повышаться в 4 раза. Хотя электрохимические измерения на клетках солевых желез не проводились, но можно, с достаточной степенью уверенности сказать, что накопление солей – активный процесс. Почти все элементы могут также транспортироваться из листьев, достигших спелости и начинающих стареть, в клетки других тканей, а также в сосуды ксилемы. Отток элементов из листьев зависит от их способности проникать во флоэму и перемещаться по ней. Показано, что основные минеральные элементы (фосфор, калий, натрий, сера, хлор, азот, и др.) перемещаются по флоэме (см. табл. 5.3). Из всех двухвалентных ионов в значительном количестве перемещается по флоэме только магний. Микроэлементы (железо, марганец, цинк, молибден) в небольшой степени перемещаются по флоэме из спелых тканей в неспелые. Поскольку железо при внутриклеточном рН нерастворимо, то оно, вероятно, перемещается в неионной форме или в виде хелатного комплекса. Многие другие элементы также образуют комплексы. Для того чтобы элементы могли переходить из сосудов ксилемы во флоэму и наоборот, существуют так называемые переходные клетки (клетки-спутники). В этих клетках на поверхности клеточных оболочек, которые находятся со стороны цитоплазмы, образуются выступы и разветвленные гребенчатые выросты. Из особенностей этих клеток вытекают два вывода, которые можно связать с транспортными функциями. Первый связан с тем, что площадь поверхности плазмалеммы в таких клетках увеличивается, так как она повторяет все изгибы оболочки. Второй вывод связан с глубоким проникновением выростов клеточной оболочки в цитоплазму, благодаря этому любая часть плазмалеммы находится вблизи клеточных органелл (ЭР, митохондрии). Эти переходные клетки бывают двух типов: А-тип служит мостиком между ксилемой и флоэмой, В-тип ответственен за перенос субстратов, транспортируемых по флоэме, в симпласт паренхимы или в клетки обкладки проводящего пучка (рис. 5.21).
Тип циркуляции ионов и метаболитов в большой мере зависит от роста и метаболической активности. Рост любого органа растения порождает потребность в растворенных веществах, обеспечивающих его метаболическую активность и поддерживающих осмотический потенциал при увеличении его объема. Таким образом, скорость роста растения влияет и на скорость поступления ионов и на распределении ионов в данной части растения. Рост растений регулируется целым рядом взаимодействующих факторов. Мы мало знаем не столько о самих факторах, сколько об их взаимодействии. Известно, что транспорт ионов регулируется гормонами. В этом случае нужно решить: имеет ли место прямое влияние гормона, обуславливающее полярный транспорт, или же это влияние роста, вызванного гормонами. ИУК влияет на структуру и вязкость мембран, а также на ионный транспорт в растительных клетках. В определенных условиях ИУК избирательно действует на поступление анионов и катионов. Наиболее интересны данные, касающиеся существования ИУК – регулируемого АТФ-зависимого механизма транспорта протонов и его связи с процессом роста клеток растяжением. ИУК в этом случае действует как активатор протонного насоса, локализованного на плазмалемме. Кинетин и другие цитокинины обладают способностью замедлять старение тканей и поддерживать нормальную скорость белкового синтеза и других метаболических процессов. Главный эффект данных регуляторов роста сводится к установлению в листе градиента метаболической активности, что сильно влияет на характер перемещения растворенных веществ. Абсцизовая кислота – гормон «старения» растений, который приводит к опаданию листьев у деревьев, обладает регуляторным действием на мембранные транспортные процессы. Она влияет на проницаемость мембран клеток корня для воды, изменяет K+/Na+ избирательность. АБК ингибирует поступление K+ в замыкающие клетки, которые необходимо для открывания устьиц. Многие транспортные процессы связаны также с фитохромной системой. Фитохром – билипротеид, существующих в двух формах (состояниях): Ф660 (Фк) и Ф730 (Фдк). Ф730 – активная форма, ответственная за включение большого числа реакций, называемых фотоморфизмами. Флоэмный транспорт. Если по ксилеме перемещение происходит исключительно от корней к листьям, то по флоэме отток наблюдается от донора (источника синтезируемых продуктов) вверх или вниз к любой точке, которая называется акцептором, т. е. местом, где продукты фотосинтеза потребляются или запасаются. Транспорт по флоэме может проходить одновременно в двух направлениях. Эта видимая двунаправленность является результатом одностороннего тока в отдельных, но смежных ситовидных трубках, которые связаны с разными донорами и акцепторами. Перед тем, как перейти к транспорту веществ по флоэме, необходимо несколько слов сказать о структуре ситовидных трубок (рис 5.22).
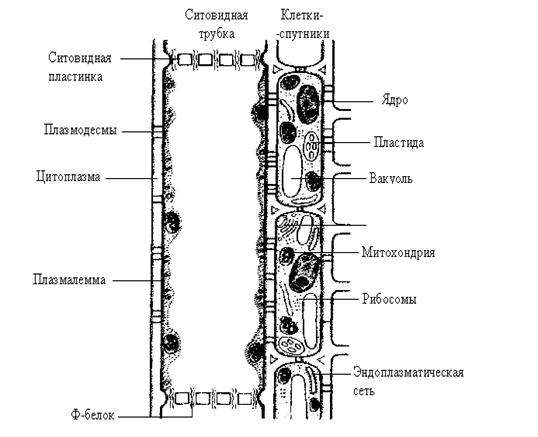
Это тонкостенные продолговатые клетки, связанные своими концами и образующие непрерывную трубу. В местах соприкосновения клеток их стенки пронизаны многочисленными ситовидными порами. Поэтому торцевые стенки называют ситовидными пластинами. Клетки ситовидных трубок в отличие от клеток ксилемных сосудов живые, хотя и отличаются от типичных живых клеток. Они не имеют ядра, но содержат некоторые другие органеллы и плазмалемму, которая окружает внешнюю поверхность клетки и ситовидные поры. О функциональной активности свидетельствует то, что клетки ситовидных трубок могут быть плазмолизированы. Тонопласт, как и ядро, и многие другие органеллы, отсутствует. В ситовидных элементах найдены белковые микрофиламенты, которые называют Ф-белками (Ф-флоэма). Структура и функция их точно неизвестны. Ситовидные трубки, как мы уже говорили, окружают клетки-спутники (переходные). Количество плазмодесм в этих клетках в 3–10 раз выше, чем в оболочках соседних клеток мезофилла листа. Есть много оснований рассматривать транспорт ассимилятов, как особый случай симпластического транспорта. О том, что ситовидные трубки принадлежат симпласту, говорят многочисленные плазмодесмы, которые соединяют ситовидные трубки с примыкающими клетками-спутниками. Среди транспортируемых органических веществ превалируют сахара. В сосудистых растениях образованные при фотосинтезе углеводы обычно перемещаются из листьев в другие части растения, например, в корни, точки роста, формирующие плоды и т. д. Дальний транспорт ассимилятов обычно идет по флоэме, но у листопадных видов древесных растений сахара могут составлять значительную часть и ксилемного сока (например, ранней весной у березы). Необходимо отметить, что глюкоза и фруктоза отсутствуют во флоэме, или присутствуют в очень малых количествах. Транспортные формы сахаров не потребляются при движении и не преобразуются в крахмал. Одним из доказательств является, как указывалось, отсутствие в проводящих сосудах инвертазы, фермента метаболизации сахарозы. Содержание аминокислот и амидов во флоэмном соке тоже регулируется далеко действующей системой «донар-акцептор». Превалируют глутамин, глутамат, аспарагин, аспартат и серин, как и в случае сахаров, их содержание то возрастает, то уменьшается. Есть данные, что некоторые аминокислоты легко поглощаются клетками высших растений, хотя эти представления требуют в дальнейшем уточнения; считают, что аминокислоты передвигаются по флоэме и перемещаются из нее в прилегающие ткани и наоборот. В эскудате флоэмы найдены и белки, в том числе имеющие ферментативную активность, нуклеиновые кислоты, аденозинфосфаты, некоторые витамины. Перемещение белков, нуклеиновых кислот и нуклеотидов маловероятно, поскольку они слабо двигаются в системе ситовидные трубки – ткани акцептора. Во флоэмном соке найдено множество органических кислот, хотя их содержание мало. Перемещение разных органических кислот на короткие и средние расстояния, несомненно, идет через мембраны или через плазмодесмы. Фитогормоны, регуляторы роста, могут перемещаться по растениям. Цитокинины и гиббереллины образуются в корнях, но, вероятно, легко перемещаются по растению, их находят в эскудатах ксилемы и флоэмы. Абсцизовая кислота, ауксин (ИУК) образуются в побеге, но могут перемещаться в корень, оба соединения найдены во флоэмном соке. В растениях перемещаются множество других природных и синтетических веществ. Ауксиноподобные соединения (α-нафтилуксусная, 2,4-дихлорфеноксиуксусная кислоты и т. д.), вероятно, перемещаются как ИУК. Скорость перемещения ассимилятов по ситовидным трубкам составляет 50–100 см/час. Перемещение веществ (ассимилятов) по ситовидным трубкам включает в себе три взаимосвязанных процесса: загрузку флоэмы, транспорт по ситовидным элементам и разгрузку флоэмы. Загрузка флоэмы происходит в результате работы Н+-помпы. А как происходит дальнейшее перемещение веществ по флоэме? Для описания механизма транспорта по ситовидным трубкам наиболее широко используется гипотеза движения под давлением, которую выдвинул в 1926 г. Мюнх. В соответствии с этой гипотезой фотосинтезирующие клетки листа образуют источник углеводов в симпласте, а ткани, использующие ассимиляты, или запасающие их (например, в виде крахмала), образуют емкость для стока. Транспорт на далекие расстояния от источника к стоку приводится в действие осмотическим градиентом давления, например между листом и корнем. Система трубок во флоэме служит каналом транспорта на далекие расстояния, в котором движется поток сока, который тянет за собой ассимиляты. Для иллюстрации такого механизма можно рассмотреть физическую систему, основанную на обычных принципах осмоса (рис. 5.23). Два сосуда с избирательно проницаемыми мембранами помещают в воду и соединяют непроницаемой трубкой (рис. 5.23, б). Сначала один сосуд заполняется концентрированным раствором сахарозы (А), а другой – разбавленным (В). В результате осмоса вода поступает в оба сосуда и в системе возникает давление. Поскольку давление значительнее увеличивается в более концентрированном растворе А, то оно будет передаваться по трубке к менее концентрированному раствору В. Так как вода поступает из А в В, то в том же направлении будет происходить и движение сахарозы.
Это движение сахарозы будет происходить до тех пор, пока концентрации растворов не уравняются. Однако, если доставлять сахарозу в А и удалять из В, то переход сахарозы из А в В будет продолжаться. Когда вода, которая удаляется из В, способна будет поступать обратно в А (соединить еще одной трубкой), то возникает постоянно работающая система. Существуют и другие гипотезы. Например, считают, что движущей силой перемещения флоэмной жидкости может быть транспорт K+ за счет электроосмотического процесса. Высказывается и гипотеза об участии фибрилл актинподобного Ф-белка пор, который взаимодействует с миозином, и таким образом, осуществляется перемещение жидкости по флоэме. Таким образом, для разгрузки флоэмы существенное значение имеет повышенное гидростатическое давление в ситовидных трубках. Кроме того, растущие и запасающие органы владеют способностью поглощать ассимиляты из флоэмы. В плазмалемме клеток акцепторной зоны функционирует Н+-помпа, которая способствует поступлению сахаров в акцепторные ткани. Сахароза поступает с участием мембранных переносчиков в симпорте с протоном, а ионы K+ – по электрохимическому градиенту. Основные различия потоков веществ, перемещаемых по флоэме и ксилеме, можно свести к следующим: – по ксилеме не переносятся сахароза и другие сахара (исключение – зимний и весенний транспорт у деревьев или у кукурузы после выметывания пестичных столбиков); – по флоэме, как правило, не переносятся ионы нитрата и сульфата; – концентрация фосфора, органического азота (аминного и амидного) и многих других ионов во флоэмном соке выше, чем в ксилемном (исключение составляет кальций); – флоэмный сок имеет слабощелочную реакцию, и уровень его рН колеблется незначительно, ксилемный сок кислый; – скорость флоэмного потока значительно ниже ксилемного. Например, максимальная скорость потока по ксилеме черешка белого люпина составляет 147 см/ч, а по флоэме 22 см/ч. При этом отдельные минеральные элементы по скорости перемещения сильно различаются и могут быть разделены на три группы: с высокой (калий, магний, фосфор, сера, азот, хлор, натрий), низкой (железо, цинк, медь, бор, молибден) и очень низкой (кальций, марганец) подвижностью.
Дата добавления: 2014-12-11 | Просмотры: 1452 | Нарушение авторских прав |