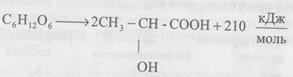|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Перетравлення і всмоктування вуглеводівРОЗДІЛ VІІ
ОБМІН ВУГЛЕВОДІВ
Перетравлення і всмоктування вуглеводів Перетравлювання вуглеводів починається у ротовій порожнині під впливом амілази слини. Їжа, змочена слиною, ковтається і потрапляє в шлунок. Він не містить ферментів, які розчеплюють вуглеводи. В шлунку α-амілаза інактивується завдяки різко кислотному середовищу (рН=1,5-2,5). Однак, у більш глибоких шарах харчової грудки, куди не відразу проникає шлунковий сік, дія слинної амілази деякий час продовжується, і полісахариди частково гідролізують до декстринів і мальтози. З шлунка їжа потрапляє у 12-и палу кишку, якраз там виникають оптимальні умови для розчеплення вуглеводів. Тут середовище слаболужне і під дією α-амілази підшлункового соку нерозчеплені полісахариди розпадаються на декстрини, а потім на мальтозу. Остання є лише проміжним продуктом, так як під впливом фермента мальтази (α-глюкозидази) гідролізує на дві молекули глюкози. Кишковий сік містить також активну сахаразу, яка гідролізує сахарозу до глюкози і фруктози, а лактаза кишкового соку розчеплює лактозу молока на глюкозу і галактозу (мал. 24). Клітковина, що міститься у харчових продуктах, в організмі людини майже не розчеплюється у зв’язку з відсутністю у травних соках ферментів, які її гідролізують. Однак, клітковина помітна входити до харчового раціону. Вона стимулює перистальтику і секрецію травних соків, сприяючи таким чином процесу і травлення. Целюлоза виводиться з організму, беручи участь у формуванні калових мас. Таким чином вуглеводи їжі розпадаються до моносахаридів, з яких складаються, і через стінки тонкого кишечника всмоктуються у кров, потрапляючи по воротній вені у печінку. Швидкість всмоктування окремих моносахаридів відрізняється, хоч молекулярна маса всіх гексоз однакова. Виявлено, що глюкоза і галактоза всмоктуються швидше, ніж інші моносахариди. Всмоктування вуглеводів являє собою складний біохімічний процес транспорту моносахаридів через мембрани клітин тонкої кишки. Виявилось важливе значення білків-переносників у цьому процесі, певну роль відіграє обмін електролітів (іонів Nа), система енергозабезпечення. Моносахариди крові використовуються для енергетичних потреб організму (70%), частково для пластичних процесів (біосинтез ліпідів, антитіл, мукополісахаридів). При фільтрації крові у нирках моносахариди реабсорбуються (всмоктуються назад у кров). У печінці моносахариди витрачаються на синтез глікогену, який тут же депонується. Запас його в організмі людини біля 350 г. Найбільше глікогену у печінці (2-5%) і в м’язах (0,5-2%). Він є легко мобілізованою полімерною формою глюкози, при її надлишку відкладається про запас, при недостачі - розчеплюється. Каталізує розпад глікогену фосфорилаза. Проміжний обмін - це сукупність усіх перетворень вуглеводів в органах і тканинах аж до утворення кінцевих продуктів. Окислення вуглеводів у тканинах є саме тою найголовнішою ланкою обміну речовин, яка забезпечує організм енергією, постачає пластичний матеріал для численних синтетичних процесів і веде до утворення кінцевих продуктів обміну - води і диоксиду вуглецю IV (СO2). Анаеробне розчеплення вуглеводів. Це розпад вуглеводів при дефіциті кисню. Якщо у такий спосіб розпадається глюкоза, то процес називають глікозом, якщо глікоген - глікогенолізом. Механізм анаеробного розчеплення вуглеводів тепер досконало вивчений. Після відщеплення від глікогену одного залишку глюкози у вигляді глюко-1-фосфату, глікогеноліз та гліколіз проходять однаково. Більша частина глікогену розпадається шляхом фосфоролізу:
n глікоген + Н3Р04
Гліколіз каталізується 11 ферментами, більшість з них виділена у чистому вигляді. Послідовність реакції гліколізу прослідкуйте за схемою 13. І. Першою ферментативною реакцією гліколізу є фосфорилювання глюкози за допомогою АТФ. Суть цього процесу полягає у перенесенні залишку фосфатної кислоти на глюкозу, реакція каталізується ферментом гексокіназою. Перша реакція гліколізу незворотна.
II. Друга реакція гліколізу - це перетворення глюкозо-6-фосфату під дією ферменту гексозофосфатізомерази у фруктозо-6-фосфат: глюкозо – 6 – Ця реакція легко протікає у двох напрямках. III. На третьому етапі фруктозо-6-фосфат знову фосфорилюється АТФ. Реакція каталізується ферментом фосфофруктокіназою:
Дана реакція проходить у присутності іонів Mg2+ і відбувається дуже повільно. Фактично, вона визначає швидкість гліколізу в цілому. IV. Четверта реакція гліколізу каталізується ферментом альдолазою і зводиться до дихотомічного поділу (грец. Dicha - на дві частини; tome - сікти). З фруктозо-1,6-дифосфату утворюються дві фосфотріози:
глюкозо – 1,6 –
V. П’ята реакція зводиться до ізомеризації тріозофосфатів, вона каталізується ферментом тріозофосфатізомеразою:
дигідроксиацетон – Четверта і п’ята реакції зворотні. Утворенням 3-фосфогліцеринового альдегіду завершується перша стадія глікозу. Друга стадія - найбільш складна і важлива, вона включає окисно-відновну реакцію і процеси фосфорилювання (синтез АТФ). VI. У шостій реакції 3-фосфогліцериновий альдегід під впливом специфічної дегідрогенази, коферменту НАД і неорганічного фосфору окислюється з утворенням 1,3-дифосфогліцеринової кислоти і відновленої форми НАД (НАДН2). Схематично цю реакцію можна зобразити так:
1,3-дифосфогліцеринова кислота являє собою макроергічну сполуку. VII. У сьомій реакції, яка каталізується фосфогліцераткіназою, відбувається передача фосфатного залишку на АДФ з утворенням АТФ і 3-фосфогліцеринової кислоти:
Таким чином, завдяки дії двох ферментів енергія, яка вивільняється при окисленні альдегідної групи 3-фосфогліцеринового альдегіду до карбоксильної групи 3-фосфогліцеринової кислоти, запасається у формі АТФ. VIII. У восьмій реакції проходить внутрішньо-молекулярне перенесення фосфатної групи і 3-фосфогліцеринова кислота перетворюється у 2-фосфогліцеринову кислоту. Реакція зворотна, протікає у присутності іонів Mg2+:
IX. У дев’ятій реакції 2-фосфогліцеринова кислота в результаті дегідратації (відщеплення Н2O) переходить у фосфоенолпіровиноградну кислоту (фосфоенолпіруват). Реакція каталізується ферментом енолазою:
Енолаза активується катіонами Мg2+, Мn2+ і гальмується F-. X. В десятій реакції проходить розрив макроергічного зв’язку і перенесення фосфатного залишку від фосфоенолпіровиноградної кислоти на АДФ. Каталізується реакція ферментом піруваткіназою:
фосфоенолпіровиноградна кислота + АДФ
XI. В одинадцятій реакції в результаті відновлення піровиноградної кислоти утворюється молочна кислота. Реакція проходить при участі фермента лактатдегідрогенази і кофермента НАДН2:
Таким чином, підводячи підсумок гліколізу необхідно відзначити, що розчеплення однієї молекули глюкози на дві молекули лактату супроводжується енергетичною ефективністю дві молекули АТФ. При глікогенолізі енергетична ефективність три молекули АТФ. Гліколіз з енергетичної точки зору малоефективний. Сумарно хімізм процесу анаеробного розчеплення глюкози можна виразити таким рівнянням:
З цієї кількості енергії близько 126 кДж розсіюється у вигляді тепла, а 84 кДж накопичується у макроергічних зв’язках АТФ. Один макроергічний зв’язок наближено містить 33,6-42,0 кДж/моль. Таким чином стає зрозумілим, звідки беруться 2 молекули АТФ при гліколізі (84:42=2). Значна частина енергії зберігається у кінцевому продукті гліколізу, в молочній кислоті. Анаеробний процес активно проходить у м’язах, при фізичних навантаженнях, внаслідок гліколізу накопичується лактат, що сприяє втомі м’язів. Далі молочна кислота потрапляє у кров, а з нею у печінку, де активно проходить її метаболізм, у тому числі і глюконеогенез (синтез глюкози з невуглеводної сировини). Біологічне значення процесу гліколізу, перш за все, полягає у вивільненні енергії, яка може певний час забезпечувати фізіологічні функції організму в умовах дефіциту кисню. Дата добавления: 2015-11-25 | Просмотры: 2103 | Нарушение авторских прав |
 глюкзозо -1 - фосфат + n — 1 глікоген
глюкзозо -1 - фосфат + n — 1 глікоген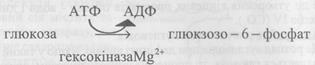
 фруктозо – 6 – фосфат
фруктозо – 6 – фосфат  фосфат
фосфат
 3 – фосфогліцери – дигідрокси –фосфат
3 – фосфогліцери – дигідрокси –фосфат  новий альдегід + ацетонофосфат
новий альдегід + ацетонофосфат новий альдегід
новий альдегід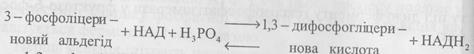



 піровиноградна кислота + АТФ
піровиноградна кислота + АТФ