|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
МЕХАНИКА МЫШЦЫМногие механические характеристики сокращающейся мышцы были изучены в первой половине XX в., до того как стала понятна сущность самого процесса сокращения на субклеточном уровне. Рассмотрим эти ставшие классическими данные с точки зрения современных знаний о лежащих в их основе механизмах. Сокращение мышцы может происходить в двух режимах: изотоническом и изометрическом. В изотоническом режиме мышца укорачивается при неизменном внутреннем напряжении, что, например, бывает при небольшой величине поднимаемого груза или сокращении мышцы без груза. В этом случае сокращение оценивают в единицах длины. При изометрическом режиме длина мышцы не меняется, а развивается внутреннее напряжение, что бывает при нагрузке мышцы неподъемным грузом. Хотя в этом режиме заметного внешнего укорочения нет, небольшое внутреннее укорочение все-таки происходит. Оно появляется за счет растягивания внутри- и внеклеточных упругих компонентов, таких, как тонкие тяжи поперечных мостиков и соединительной ткани, располагающихся последовательно с мышечными волокнами. Сокращение при изометрическом режиме оценивают в единицах силы. Необходимо указать на одно чрезвычайно важное положение о распределении напряжения в мышце. Его смысл состоит в том, что при данной нагрузке напряжение в любом элементе линейного ряда (например, в одном звене цепи) равно напряжению, развиваемому в каждом из остальных элементов этого ряда. Это положение справедливо как для активного напряжения, вызываемого сокращением, так и для пассивного напряжения, обусловленного натяжением мышцы под действием внешнего груза. Скорость, с которой мышца укорачивается при данной нагрузке, линейно зависит от числа последовательных саркомеров в мышце. Допуская, что все саркомеры одного мышечного волокна укорачиваются в единицу времени на одну и ту же величину, общее уменьшение длины за определенное время будет пропорционально числу последовательно соединенных саркомеров. Отсюда следует, что противоположные концы длинной мышцы сближаются с большей скоростью, чем короткой. Соответственно из двух мышц одинаковой длины сокращение будет происходить быстрее у той мышцы, у которой более короткие саркомеры. При этом скорость скольжения между филаментами будет одинаковой в обеих мышцах. Вместе с тем максимальная скорость сокращения и максимально развиваемая сила мышцы будут находиться в обратной зависимости от длины саркомера. Так, у мышцы с длинными саркомерами (т. е. большее перекрывание филаментов) будут развиваться большая сила и низкая предельная скорость укорочения, и наоборот, в мышце с короткими саркомерами сила будет меньше, но скорость больше. Величина усилия, развиваемого при сокращениях, ограничена числом миофибрилл, а точнее — числом миозиновых и актиновых филаментов, работающих параллельно. Поэтому толстая мышца способна поднять больший груз, чем тонкая, независимо от длины. Известно, что по мере увеличения нагрузки на мышцу удлиняется время (латентный период), которое мышца затрачивает на отрыв груза от поверхности опоры. Причина заключается в том, что мышце необходимо время, для того чтобы поперечные мостики, активируясь, развили усилие. Чем больше груз, тем продолжительнее время, требуемое на растяжение эластических компонентов и создание необходимого усилия (рис. 2.27). С активацией поперечных мостиков связана и зависимость скорости укорочения мышцы от нагрузки. Скорость укорочения падает по мере увеличения силы, которую должна развить мышца для поднятия груза (см. рис. 2.27). Это обусловлено склонностью актиновых филаментов «пробуксовывать» под влиянием силы, возникающей при активации поперечных мостиков. В функциональном отношении мышцу можно представить как сократительный компонент, соединенный параллельно с первым эластическим компонентом (сарколемма, соединительная ткань и т.д.) и последовательно со вторым эластическим компонентом (рис. 2.28). В последовательно включенный компо-
А — начало сокращения. Мышца представлена как сократительный элемент, соединенный последовательно и параллельно с эластическим элементом; Б — напряжение в мышце нарастает по мере растягивания последовательно расположенных эластических элементов; до определенного момента длина мышцы неизменна; В — при равенстве механического напряжения в мышце и массы груза последний падает и длина мышцы уменьшается (сокращение)
нент входят сухожилия, соединительная ткань, прикрепленная к сухожилиям. Согласно закону Гука длина объекта с идеальной эластичностью увеличивается пропорционально приложенной к нему силе. На схеме эластические элементы представлены в виде пружин. Укорочение или напряжение мышцы достигает своего максимума в течение 10...500 мс и зависит от типа мышцы, температуры и величины внешней нагрузки. Напомним, что при возбуждении и выходе ионов кальция из саркоплазматического ретикулума поперечные мостики сначала прикрепляются к актиновым нитям, а затем начинается процесс активного скольжения. Причем согласно строению мышцы активное скольжение сначала должно вызвать растяжение последовательного эластического элемента, прежде чем начнет развиваться укорочение всей мышцы (см. рис. 2.28). В зависимости от частоты раздражения у мышц различают два вида сокращений: одиночное и тетаническое. Одиночное сокращение возникает при возбуждении мышечных волокон одним потенциалом действия. Во время одиночного сокращения прикрепление миозиновых мостиков к актиновым филаментам быстро устраняется вследствие того, что ионы кальция возвращаются в сар-коплазматический ретикулум. При этом филаменты не успевают втянуться друг в друга достаточно глубоко, чтобы растянуть ткани, входящие в последовательный эластический компонент, развив в мышце полное напряжение (рис. 2.29). Поэтому напряжение, на которое способна сократительная система, не может реализоваться в одиночном мышечном сокращении. В то же время если второй потенциал действия последует за первым прежде, чем в сар-коплазматический ретикулум возвратятся ионы кальция, активное состояние сократительных компонентов будет продлено. При генерации от трех-четырех потенциалов действия состояние изометрического напряжения мышцы продолжает нарастать до тех пор, пока напряжение, вызванное внутренним укорочением со- Время Рис. 2.29. Одиночное и тетаническое сокращения мышцы кратительных компонентов, не станет достаточным, чтобы вызвать «пробуксовку» поперечных мостиков, препятствующих дальнейшему внутреннему укорочению. Такое сокращение мышцы называется тетаническим. В зависимости от частоты следования мышечных потенциалов действия возникают разные степени слияния одиночных сокращений и тетанического напряжения (см. рис. 2.29). Источником энергии для сократительного процесса служит АТФ, что видно из выражения АТФ <^ АДФ + Р + Свободная энергия. Однако концентрация АТФ в мышце довольно низкая, поэтому при длительном сокращении мышцы АТФ должно хватить ненадолго. В то же время АТФ быстро регенерирует за счет фосфори-лирования АДФ, при этом высокоэнергетическая фосфатная группа поставляется за счет ферментативного расщепления богатого фосфором соединения — креатинфосфата. Этот процесс катализирует фермент креатинфосфокиназа: Креатинфосфат + АДФ «=* Креатин + АТФ. Данная реакция протекает достаточно энергично, поставляя АТФ для физической нагрузки. Сокращение мышц сопровождается потерей энергии в форме теплоты. Коэффициент полезного действия (КПД) мышцы как своебразной механической машины вычисляется по формуле кпд = и/гл + а)]Ю0%, где А — механическая работа; й —тепловой выход мышцы, который включает в себя теплообразование, связанное с электрическим возбуждением и механической активацией мышцы, а также теплоту сокращения, пропорциональную величине укорочения мышцы.
1 моль АТФ при распаде высвобождает 48 кДж энергии, и для ресинтеза 1 моль АТФ в организме животного необходимо 3 моль кислорода. В условиях интенсивной мышечной работы запасов кислорода не хватает для быстрого ресинтеза АТФ. В итоге в организме накапливается много недоокисленных продуктов (молочной кислоты и др.). Создается так называемая кислородная задолженность, погашаемая после работы за счет автоматического усиления дыхания и кровообращения. Если же работа, несмотря на наличие кислородного долга, продолжается, то наступает утомление мышцы. В нормальных условиях утомление животного развивается прежде всего как отказ работы структур ЦНС. Децентрализованные мышцы (т. е. при перерезке нерва) при раздражении работают дольше, однако в конечном счете также утомляются. Скорость утомления у мышц разная в зависимости от типа энергетического обеспечения мышц. Дата добавления: 2015-12-15 | Просмотры: 1079 | Нарушение авторских прав |
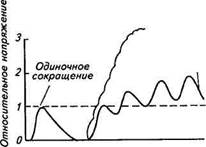
 Рис. 2.28. Механизмы сокращения мышцы:
Рис. 2.28. Механизмы сокращения мышцы: Энергия, освобождающаяся в виде теплоты, приблизительно в 5 раз превышает энергию, затраченную на выполнение механической работы. Обычно КПД мышцы не превышает 20...30 %. Однако здесь важно отметить, что мышца — не только механическая машина, но и основной обогреватель организма, поэтому тепловой выход небесполезен.
Энергия, освобождающаяся в виде теплоты, приблизительно в 5 раз превышает энергию, затраченную на выполнение механической работы. Обычно КПД мышцы не превышает 20...30 %. Однако здесь важно отметить, что мышца — не только механическая машина, но и основной обогреватель организма, поэтому тепловой выход небесполезен.