|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ПЕРЕДАЧА НЕРВНОГО ВОЗБУЖДЕНИЯ МЕЖДУ КЛЕТКАМИ. ПРЕДСТАВЛЕНИЕ О СИНАПСАХ
Мы рассмотрели механизмы передачи возбуждения в пределах одной клетки. Что же происходит, когда сигнал доходит до места контакта клетки с другой — нервной, мышечной или иной клеткой организма? Здесь целесообразно сделать небольшое историческое отступление. В начале XX в. крупнейший гистолог С. Рамон-и-Кахаль с помощью светового микроскопа выяснил, что нейроны в нервной ткани окрашиваются как отдельные клетки. Однако многие анатомы продолжали считать, что нервная система представляет собой сеть, в которой цитоплазма-тическое пространство отростков одной клетки напрямую сообщается с внутриклеточным объемом других клеток. Окончательно этот вопрос был разрешен в 40-х годах с появлением электронного микроскопа, когда были получены четкие данные о наличии мембраны вокруг нервных клеток и их окончаний, а также о существовании особых участков межнейронного взаимодействия, получивших название синапсов.
Рис. 2.19. Схема электрического синапса: стрелкой показано направление распространения потенциала действия в пресинаптическом нервном волокне
достижении порогового значения в ней начнется регенеративный процесс повышения проводимости к ионам натрия и возникнет потенциал действия, который может проследовать дальше по отросткам клетки. Такое соединение называется электрический синапс. Передача через электрический синапс электрического сигнала затруднена несовершенством кабельных свойств клеток. В частности, в том случае, если площадь контакта мембраны нервного волокна и сомы клетки сравнительно небольшая, например контактирует нервный отросток диаметром 5 мкм, общее электрическое сопротивление мембраны этого отростка и участка сомы клетки, к которому он примыкает, составляет миллиарды ом. В то же время сопротивление клеточной мембраны всей сомы клетки будет в несколько десятков раз меньше и сыграет роль короткого замыкания для тока, выходящего из концевой мембраны отростка. Кроме того, ионный ток будет утекать через пространство между мембранами. Все это, несмотря на высокий фактор надежности — 5... 6 (отношение амплитуды потенциала действия к величине пороговой деполяризации), для потенциала действия чрезвычайно сильно ослабит выходящий из отростка ионный ток и передача возбуждения в этом участке не будет происходить. Для того чтобы передача функционировала более эффективно, необходимо увеличить площадь соприкосновения двух мембран, а также плотнее приблизить сами мембраны. Непосредственная электрическая передача была экспериментально обнаружена сначала на толстых (гигантских) волокнах ракообразных, а затем такие соединения были найдены и у позвоночных животных между нервными клетками в ЦНС, в гладких и сердечных мышцах. Отметим, что электрическая передача возбуждения имеет определенные преимущества. Поскольку ионный ток при такой передаче непосредственно течет из передающей (пресинаптической) клетки в воспринимающую (постсинаптическую) клетку без каких-либо промежуточных этапов, задержка при проведении возбуждения будет минимальная. Таким образом, электрическое проведение более удобно в тех случаях, когда необходимо быстро охватить возбуждением несколько нервных клеток. Вместе с тем в большинстве синапсов нервной и нервно-мышечной систем для передачи сигналов используются специальные химические вещества-посредники — медиаторы. Поскольку следующая часть данной главы посвящена изложению механизмов возбуждения в мышцах, здесь мы остановимся на синаптическом проведении через нервно-мышечный синапс или, как часто называют, через концевую пластинку скелетных мышц позвоночных. О строении и функции синапсов в ЦНС будет подробнее изложено в гл. 12. Схематически последовательность событий, происходящих при передаче электрического сигнала через химический синапс, можно представить следующим образом (рис. 2.20). После возникно- Х^\ ПД в пресинаптическом ^----- s Связывание медиатора с \fj,tj волокне \) постсинаптическими рецепто- Активация синаптических каналов;
\\_^^ Высвобождение медиатора ч>\^ ПД в постсинаптической Рис. 2.20. Механизм передачи электрического сигнала через химический синапс вения потенциала действия в пресинаптическом окончании мембрана его деполяризована. При этом помимо натриевых и калиевых каналов активируются (открываются) кальциевые каналы и в окончание из внешней среды входят ионы кальция. Повышение внутриклеточной концентрации кальция вызывает экзоцитоз везикул, наполненных медиатором (выброс из них медиатора). Содержимое везикул поступает во внеклеточное пространство, и часть молекул медиатора, диффундируя к постсинаптической мембране (мембрана мышечной клетки), связывается со специальными (рецепторными) участками этой мембраны. В результате происходит активация ее ионных каналов, связанных с этими молекулами. Движение по каналам соответствующих ионов по их электрохимическим градиентам порождает постсинаптичес-кий ток, под действием которого возникает деполяризующий постсинаптический потенциал мышечного волокна. Если этот потенциал превышает пороговый уровень, возникает потенциал действия. Из-за наличия химического звена проведение в химическом синапсе происходит медленнее, чем в электрическом. Однако химическая передача гибче электрической, поскольку с ее помощью легко может осуществляться как возбуждающее, так и тормозное действие. Кроме того, при активации постсинаптичес-ких каналов химическими агентами может возникать достаточно сильный ток, способный деполяризовать до порогового уровня крупные клетки. В связи с этим при химической передаче тонкие пресинаптические волокна могут возбуждать большие постсинап-тические клетки. Необходимо отметить, что принятию представленной схемы химической передачи предшествовали в течение шести десятиле-
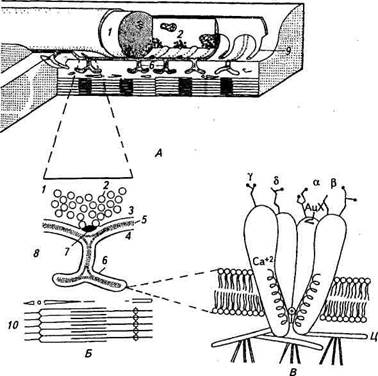
Вернемся вновь к нервно-мышечному синапсу — концевой пластинке. На рисунке 2.21, А представлено трехмерное изображение концевой пластинки. Она состоит из специализированных участков постсинаптической мембраны, являющейся частью мембраны мышечного волокна, окончания нервного волокна (окончания двигательного нерва — пресинаптической мембраны) и шванновских клеток. От окончания нерва отходят веточки толщиной около 2 мкм, каждая из которых лежит в продольном углублении мембраны мышечного волокна. Мембрана, выстилающая это углубление, с периодичностью 1...2 мкм образует поперечные субнейрональные складки. В участках нервного окончания, расположенного непосредственно над этими складками, имеются так называемые активные зоны — поперечные участки с несколько утолщенной пресинаптической мембраной, над которыми скапливаются синаптические пузырьки. Пузырьки выделяются из активных зон путем экзоцитоза. Выделение медиатора из пресинаптического окончания начинается под действием деполяризации этого окончания, вызванной потенциалом действия. Высвободившийся медиатор, в данном случае это АцХ, связывается с рецептором, т. е. со специальной белковой молекулой в постсинаптической мембране концевой пластинки, в результате чего на короткое время открываются каналы, через которые течет ионный ток. Для выяснения природы ионов, обусловливающих синаптичес-кий ток, первоначально изменяли внеклеточные концентрации различных ионов и исследовали влияние этих изменений. Так, было обнаружено, что входящий синаптический ток в концевой пластинке является натриевым ионным током, причем он частично компенсируется выходящим калиевым током. Дальнейшие исследования показали, что через одни и те же каналы, активируемые в концевой пластинке АцХ, проходят ионы натрия и калия. Значит, эти каналы обладают меньшей избирательностью (селективностью), чем потенциалзависимые натриевые и калиевые каналы при деполяризации мембраны. Мы уже подробно разбирали механизмы формирования мембранного потенциала в нервных и мышечных клетках и знаем, что
Рис. 2.21. Схема нервно-мышечного синапса (концевой пластинки) и ацетил-холинактивируемого ионного канала: А - тоехмеоная реконструкция нервно-мышечного синапса: 1 - терминальная часть нервного при изменении мембранной проницаемости для какого-либо иона мембранный потенциал согласно уравнению Голдмана (17) оудет смещаться до нового уровня. Если проницаемость для данного иона увеличится, мембранный потенциал сместится ближе к равновесному потенциалу для этого иона. И наоборот, уменьшение проницаемости вызовет смещение в противоположную сторону от равновесного потенциала. В ответ на АцХ каналы концевой пластинки становятся одновременно проницаемы для ионов натрия и калия. При этом возникает смешанный ионный ток, под деистви- ем которого потенциал постсинаптической мембраны смещается к новому уровню, так называемому потенциалу реверсии, занимающему промежуточное положение между равновесными потенциалами для этих двух проникающих ионов и равному -10 мВ. В общем случае потенциал реверсии для ионных токов, переносимых двумя ионами, зависит от двух главных факторов: относительной проницаемости активированного канала для проникающих ионов; равновесных потенциалов для этих ионов, зависящих, в свою очередь, от их концентрационных градиентов. Постсинаптические каналы открываются на короткое время (на несколько мс), поскольку АцХ быстро подвергается ферментативному расщеплению ацетилхолинэстеразой (АцХЭ), которая располагается в области субнейрональных складок. АцХ- активируемый канал. Успехи в совершенствовании методов молекулярной биологии, а также электрофизиологических методов позволили выделить, идентифицировать и изучить АцХ-ак-тивируемый канал — интегральный белок мышечной мембраны. АцХ-активируемый канал состоит из пяти белковых субъединиц, которые образуют трубчатую структуру с общей молекулярной массой около 250 000. Канал выступает из мембраны с обеих сторон, причем над наружной ее поверхностью возвышается воронкообразная структура — вход в канал (см. рис. 2.21, В). Рецептор-ный участок канала состоит из двух белковых субъединиц и располагается на наружной поверхности мембраны. Каждая из белковых субъединиц содержит участок связывания АцХ. Если к этим двум участкам присоединяется АцХ, то канал с высокой вероятностью переходит из закрытого состояния в открытое и через него течет ионный ток. Регистрацию ионного тока через отдельные АцХ-каналы удалось осуществить с помощью методики локальной фиксации немецкими учеными Э. Неером и Б. Сакманом в середине 70-х годов XX в. (см. функционирование потенциалзависимого натриевого канала). Здесь надо отметить, что в норме АцХ-каналы и связанные с ними АцХ-рецепторы сосредоточены в постсинаптической мембране концевой пластинки и плотность их весьма велика — около 104/мкм2. В остальной части мышечной мембраны их нет, и именно из-за этого в норме не иннервированная часть мышечных волокон нечувствительна к действию АцХ. Однако если перерезать двигательный нерв, то зона чувствительности к АцХ постепенно распространяется от концевой пластинки, охватывая всю мембрану мышечного волокна. Значит, рецепторы и каналы появляются во внесинаптических участках. В нормальных условиях эти внесинаптические каналы каким-то образом блокированы малоизученным трофическим влиянием нервной клетки, иннерви-рующей мышечное волокно. Таким «рассеянным» распределением АцХ-каналов воспользовались Неер и Сакман. На участок мембраны накладывали пипетку и с помощью небольшого вакуума плотно присасывали ее к мембране. Микропипетка была заполнена физиологическим раствором, содержащим АцХ или его агонист (т. е. вещество, оказывающее сходное с АцХ действие). С целью увеличения ЭДС для входящего тока мембранный потенциал сдвигали в сторону гиперполяризации. Усилитель регистрировал очень малые, менее 5 • 10~^2 А, входящие кратковременные токи, которые по форме сходны с токами одиночных потенциалзависимых натриевых каналов, т. е. имеют форму прямоугольных импульсов и подчиняются закону «все или ничего». Частота таких токов была пропорциональна концентрации АцХ или его агониста в растворе пипетки. Если же эти вещества отсутствовали, то токи не регистрировались. Проводимость одиночного АцХ-канала крайне мала и составляет около 20пкС, что соответствует 5 • 10'4 Ом, т. е. того же порядка, что и у потенциалзависимых натриевых каналов. Статистический анализ одиночных токов показал, что каналы случайным образом изменяют свое состояние (флуктуируют), переходя из закрытого состояния в открытое. По-видимому, связывание молекулы АцХ с рецептором закрытого канала резко увеличивает вероятность его открывания, когда ионы в течение короткого времени могут проходить через данный канал. Дата добавления: 2015-12-15 | Просмотры: 899 | Нарушение авторских прав |

 Прежде чем перейти к описанию структуры и функции синапсов, рассмотрим ситуацию, когда замкнутая клеточная мембрана нервного отростка примыкает к мембране сомы другой нервной клетки или к мембране мышечного волокна. Допустим, в мембране конца нервного отростка (аксона) в результате распространения возбуждения возник потенциал действия. Тогда согласно вышеописанной схеме (см. рис. 2.17) от этого участка через его мембрану и мембрану сомы другой клетки будут пассивно распространяться петли тока (рис. 2.19), деполяризуя мембрану сомы соседней клетки. При
Прежде чем перейти к описанию структуры и функции синапсов, рассмотрим ситуацию, когда замкнутая клеточная мембрана нервного отростка примыкает к мембране сомы другой нервной клетки или к мембране мышечного волокна. Допустим, в мембране конца нервного отростка (аксона) в результате распространения возбуждения возник потенциал действия. Тогда согласно вышеописанной схеме (см. рис. 2.17) от этого участка через его мембрану и мембрану сомы другой клетки будут пассивно распространяться петли тока (рис. 2.19), деполяризуя мембрану сомы соседней клетки. При V-/, Увеличение проницаемости Созывающего лостсиналтический
V-/, Увеличение проницаемости Созывающего лостсиналтический
 тий XX в. споры среди ученых о наличии химической передачи и медиаторов в нервно-мышечной системе организма. Первые прямые данные в пользу существования химического медиатора были получены в 20-е годы XX в. О. Леви. Он обнаружил, что при торможении работы сердца лягушки путем раздражения блуждающего нерва из сердца в кровь выделяется вещество, под действием которого частота сокращений сердца другой лягушки также уменьшается. В последующих работах было установлено, что этим веществом — медиатором является ацетилхолин (АцХ). В дальнейшем был обнаружен ряд других медиаторов, накоплено множество данных об их действии и химической природе.
тий XX в. споры среди ученых о наличии химической передачи и медиаторов в нервно-мышечной системе организма. Первые прямые данные в пользу существования химического медиатора были получены в 20-е годы XX в. О. Леви. Он обнаружил, что при торможении работы сердца лягушки путем раздражения блуждающего нерва из сердца в кровь выделяется вещество, под действием которого частота сокращений сердца другой лягушки также уменьшается. В последующих работах было установлено, что этим веществом — медиатором является ацетилхолин (АцХ). В дальнейшем был обнаружен ряд других медиаторов, накоплено множество данных об их действии и химической природе.