|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Головні властивості проведення збудження через синапси наступні.1. Однобiчнiсть проведення збудження. В аксонi збудження проходить в обох напрямках вiд мiсця його виникнення, у нервовому центрi – тiльки в одному напрямку: вiд рецептора до ефектора (тобто на рівні синапса від пресинаптичної мембрани до постсинаптичної), що пояснюється структурно-функціональною органiзацiєю синапса. 2. Синаптична затримка проведення збудження. Збудження у нервовому центрi проводиться з меншою швидкiстю, нiж в iнших частинах рефлекторної дуги. Це пов'язано з часом, що витрачається на процеси видiлення медiатора, з фізико- хiмiчними процесами, якi вiдбуваються в синапсi, з виникненням ЗПСП i генерацiєю ПД. На все це в одному синапсi витрачається 1,5-2 мс. Таке явище дiстало назву синаптичної затримки проведення збудження. Чим складнiша рефлекторна дуга, тим бiльше синапсiв i, вiдповiдно, бiльша синаптична затримка. Сума синаптичних затримок у рефлекторній дузі отримала назву справжнього часу рефлексу. Час вiд початку дiї подразника до прояви рефлекторної вiдповiдi називається прихованим, або латентним перiодом (ЛП) рефлексу. Тривалiсть цього перiоду залежить вiд кiлькостi нейронiв, а отже й синапсів, якi беруть участь у рефлексi. Наприклад, сухожильний колiнний рефлекс, рефлекторна дуга якого моноси- наптична, має ЛП 24 мс, зорова або слухова реакцiя – 200 мс. 3. Сумацiя збуджень – явище виникнення збудження при певних умовах нанесення пiдпорогових под- разнень. Сумацiю описано I.М.Сєченовим. Є два види сумацiї (рис. 2.8). Часова сумацiя – виникнення збудження на ряд пiдпорогових подразнень, що послiдовно надходять до клiтини чи центру вiд одного рецепторного поля. Частота стимулiв повинна бути такою, щоб iнтервал мiж ними був не бiльше 15 мс, тобто коротшим за тривалiсть ЗПСП. При таких умовах ЗПСП на черговий стимул розвивається до того, як завершиться ЗПСП на попереднiй стимул. ЗПСП сумуються, їх амплiтуда зростає i, нарешті, при досягненнi критичного рівня деполяризації, виникає ПД. Просторова сумацiя – виникнення збудження при одночасному нанесеннi декількох допорогових стимулів на рiзнi дiлянки рецепторного поля. Якщо ЗПСП виникають одночасно в декiлькох синапсах нейрона (не менше 50), мембрана нейрона деполяризується до критичних значень i, як наслідок, виникає ПД.
А – часова сумацiя. Частота стимулiв повинна бути такою, щоб iнтервал мiж ними був не бiльше 15 мс, тобто коротшим за тривалiсть ЗПСП. При таких умовах ЗПСП на черговий стимул розвивається до того, як завершиться ЗПСП на попереднiй стимул. ЗПСП сумуються, їх амплiтуда зростає i, нарешті, при досягненнi критичного рівня деполяризації, виникає ПД. Б – просторова сумацiя. ЗПСП виникають одночасно в декiлькох синапсах нейрона (не менше 50), мембрана нейрона деполяризується до критичних значень i, як наслідок, виникає ПД.
4. Трансформацiя ритму збудження – невiдповiднiсть частоти ПД в аферентній та еферентній ланках рефлекторної дуги. Трансформацiя понижуючого типу відтворюється при високочастотному чутливому подразненні та обумовлена тривалiстю в синапсах абсолютної рефрактерної фази, пiд час якої блокується частина ПД, що надходять по аферентнiй частинi рефлекторної дуги. Трансформацiя пiдвищуючого типу пояснюється порівняно великою тривалiстю ЗПСП критичного рiвня, пiд час якого генерується декiлька ПД. У розвитку цього виду трансформацiї вiдiграють також роль структури, в яких збудження циркулює по колу, або розповсюджується по декiлькох ланцюгах з рiзною кiлькiстю синаптичних перемикань. У першому i другому випадках на еферентну частину рефлекторної дуги виходить декiлька ПД у вiдповiдь на один аферентний стимул (рис. 2.9). 5.
Рис. 2.9. Механізми розвитку трансформації пiдвищуючого типу. А - збудження циркулює по колу, Б – збудження розповсюджується по декiлькох ланцюгах з рiзною кiлькiстю синаптичних перемикань.
пам’яті. 6. Посттетанiчна потенцiацiя – явище появи або пiдсилення вiдповiдi на поодинокi тестуючi сенсорнi стимули протягом деякого часу пiсля попереднього слабкого частого (100-200 iмп/с) ритмiчного подразнення. Потенцiацiя обумовлена процесами на рівні пресинаптичної мембрани й виражається збiльшенням виділення медіатора. Це явище має гомосинап- тичну природу, тобто виникає в тому випадку, коли ритмiчне подразнення i тестуючий iмпульс надходять до нейрона по одних i тих же аферентних волокнах. В основу потенцiацiї покладено, перш за все, підсилення надходження Са2+ через пресинаптичну мембрану. Це явище прогресивно зростає з кожним імпульсом. І коли кількість Са2+ стає більшою ніж мітохондрії та ендоплазматичний ретикулюм здатні аб- сорбувати, настає пролонговане звільнення медіатора в синапс. Отже, має місце мобілізація готовності
до виділення медіатора більшою кількістю везикул і як наслідок – збільшення кількості квантів медіатора. За сучасними даними в генезі посттетанічної потенціації має значення секреція ендогенних нейропептидів, особливо при переході короткочасної потенціації в довготривалу. Серед них нейромодулятори, що діють як на пресинаптичну, так і постсинаптичну мембрани. Стимуляторами є атропін, соматостатин, фактор росту, а інгібіторами – інтерлейкін, тироліберін, мелатонін. Значимі також арахідонова кислота, NO. Потенціація має значення при організації пам’яті, зокрема, короткочасної. За рахунок підсилюючих ланцюгів організується навчання. 7. Стомлюванiсть нервових центрiв. При тривалому повторному виконаннi того ж самого рефлексу через деякий час настає стан зменшення сили рефлекторної реакцiї i навiть повне її пригнiчення, тобто настає втома. Пiд втомою розумiють зниження працездатностi, викликане працею. Втома першочергово розвивається у нервовому центрi. Втома пов'язана з порушенням передачi в синапсах, виснаженням ресурсiв медiатора у пресинаптичних везикулах, зниженням чутливостi рецепторiв субсинаптичної мем- брани до медiаторiв, а також послабленням дiї ферментних систем. Однiєю з причин є «звикання» постсинаптичної мембрани до дiї медiатора – габiтуацiя. Причина її вбачається в зниженні амплітуди ЗПСП, «віддаленні» ЗПСП від критичного рівня деполяризації. Можливе також погіршення проникливості Са2+ через пресинаптичну мембрану. Нервовi центри рiзних рефлексiв мають рiзну швидкiсть стомлення. Найменше стомлюються центри пропрiоцептивних тонiчних рефлексiв, що забезпечують пiдтримання тонусу посмугованих м'язiв. Значно швидше стомлюються центри довiльних швидких рухiв, розташовані в вищих відділах ЦНС. Процеси працездатності, втоми та здатності до відновлення вивчав І.М.Сєченов. Він встановив, що втомлена кінцівка відновлює працездатність швидше, якщо в період відпочинку друга кінцівка працює. Це так званий «активний відпочинок». І.П.Павлов запропонував для відновлення працездатності чередувати фізичну й розумову діяльність. 8. Чутливiсть нервових центрiв до нестачi кисню та хiмiчних речовин. Нервовi центри дуже чутливi до змін хiмiчного складу кровi, тканинної рiдини, дефiциту кисню. Останнє пов'язано з тим, що єдиним джерелом енергетичного забезпечення ресинтезу АТФ у ЦНС є окисне фосфорилювання. Тому нестача кисню призводить до виснаження енергетичних резервiв. Крiм того, нервовi клiтини характеризуються iнтенсивним обмiном i споживанням кисню. 1/6 – 1/8 частина кисню, що вживається організмом людини, витрачається нервовою системою. На 100 гр. тканини мозку кисню вживається в 22 рази більше, ніж на таку ж масу скелетного м'яза в стані спокою. Тому навiть короткочасна зупинка мозкового кровообiгу веде до розладу в ЦНС. Особливо страждають нейрони кори великих півкуль. Через 5-6 хвилин у них розвива- ються незворотні процеси, що призводять до загибелi. Центри мозкового стовбура менш чутливi, вони гинуть через 15-20 хвилин, спинного мозку – через 20-30 хвилин. Усе це обмежує граничні строки доцільної реанімації людини в стані клінічної смерті. Деякi хiмiчнi речовини специфiчно впливають на вiдповiднi нервовi центри, що пов'язано зі структура- ми цих хiмiчних речовин, якi можуть бути спорідненими із вiдповiдними медiаторами нервових центрiв. Особливу групу складають нейротропні речовини. Серед них: 1) наркотизуючі – такі, що використовуються в хірургічній практиці для наркозу (хлоретил, кетамін, барбітурати та ін.); 2) транквілізатори – заспокійливі засоби (реланіум, аміназин, триоксазин, амізіл, оксилідин; серед рослин- них препаратів – настій собачої кропиви, півонії та ін.); 3) нейротропні речовини вибіркової дії (лобелін, цитітон – збуджувачі дихального центру; апоморфін – збуджувач центру блювоти; мескалін – зоровий галюциноген та ін.). Досить складне i важливе питання про дiю наркотикiв на ЦНС та їх небезпечність.
Дата добавления: 2015-12-15 | Просмотры: 876 | Нарушение авторских прав |
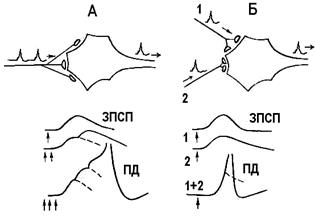 Рис. 2.8. Сумація збудження.
Рис. 2.8. Сумація збудження.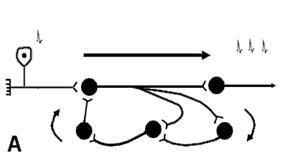 Пiслядiя збудження – явище продовження збудження в ЦНС пiсля припинення подразнення. Спричиняється механізмами, аналогічними таким при трансформації підвищуючого типу. Пiслядiя може бути короткочасною (десятки i сотнi мілісекунд) i тривалою (секунди i значно бiльше). Короткочасна пiслядiя пов'язана з великою тривалiстю ЗПСП критичного рiвня.
Пiслядiя збудження – явище продовження збудження в ЦНС пiсля припинення подразнення. Спричиняється механізмами, аналогічними таким при трансформації підвищуючого типу. Пiслядiя може бути короткочасною (десятки i сотнi мілісекунд) i тривалою (секунди i значно бiльше). Короткочасна пiслядiя пов'язана з великою тривалiстю ЗПСП критичного рiвня.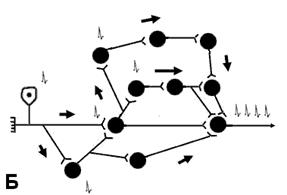 Тривала пiслядiя зумовлена циркуляцiєю збудження по замкнених нервових ланцюгах. Таке явище зветься реверберацiєю. Воно забезпечує активнiсть нервових центрiв навiть при вiдсутностi аферентних сигналiв. Зав- дяки реверберацiї збуджень (ПД) нервовi центри постiйно знаходяться в станi тонусу. Вони спонтанно посилають iмпульси до робочого органу. Тонус нерво- вих центрiв забезпечує тонус посмугованих i непосму- гованих м'язових волокон, стiнок кровоносних судин. Реверберацiя збудження в кiльцевих сiтках пiдтримується рiзними аферентацiями з боку рефлексо- генних зон iнших рефлексiв, а також рiдинним середо- вищем органiзму. Закономірності розвитку реверберації на рівні цілісного організму важливі при організації
Тривала пiслядiя зумовлена циркуляцiєю збудження по замкнених нервових ланцюгах. Таке явище зветься реверберацiєю. Воно забезпечує активнiсть нервових центрiв навiть при вiдсутностi аферентних сигналiв. Зав- дяки реверберацiї збуджень (ПД) нервовi центри постiйно знаходяться в станi тонусу. Вони спонтанно посилають iмпульси до робочого органу. Тонус нерво- вих центрiв забезпечує тонус посмугованих i непосму- гованих м'язових волокон, стiнок кровоносних судин. Реверберацiя збудження в кiльцевих сiтках пiдтримується рiзними аферентацiями з боку рефлексо- генних зон iнших рефлексiв, а також рiдинним середо- вищем органiзму. Закономірності розвитку реверберації на рівні цілісного організму важливі при організації