|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
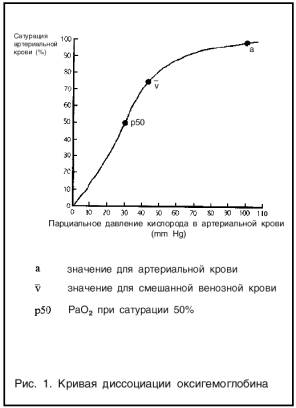
Транспорт кислорода кровью.Кислород в крови находится в растворенном виде и в соединении с гемоглобином. В плазме растворено очень небольшое количество кислорода, каждые 100 мл плазмы крови при напряжении кислорода (100 мм рт. ст.) могут переносить в растворенном состоянии лишь 0.3 мл кислорода. Это явно недостаточно для жизнедеятельности организма. При таком содержании кислорода в крови и условии его полного потребления тканями минутный объем крови в покое должен был бы составлять более 150 л/мин. Важен другой механизм переноса кислорода путем его соединения с гемоглобином. Каждый грамм гемоглобина способен связать 1.34 мл кислорода. Максимальное количество кислорода, которое может быть связано 100 мл крови, - кислородная емкость крови (18,76 мл или 19 об%). Кислородная емкость гемоглобина - величина, отражающая количество кислорода, которое может связаться с гемоглобином при его полном насыщении. Другой показатель дыхательной функции крови - содержание кислорода в крови, который отражает истинное количество кислорода, как связанного с гемоглобином, так и физически растворенного в плазме. В 100 мл артериальной крови в норме содержится 19-20 мл кислорода, в таком же объеме венозной крови — 13-15 мл кислорода, при этом артерио-венозная разница составляет 5-6 мл. Показатель степени насыщения гемоглобина кислородом – отношение количества кислорода, связанного с гемоглобином, к кислородной емкости последнего. Насыщение гемоглобина артериальной крови кислородом у здоровых лиц составляет 96%. Образование оксигемоглобина в легких и его восстановление в тканях находится в зависимости от парциального напряжения кислорода крови: при его повышении насыщение гемоглобина кислородом возрастает, при понижении — уменьшается. Эта связь носит нелинейный характер и выражается кривой диссоциации оксигемоглобина, имеющей S-образную форму.
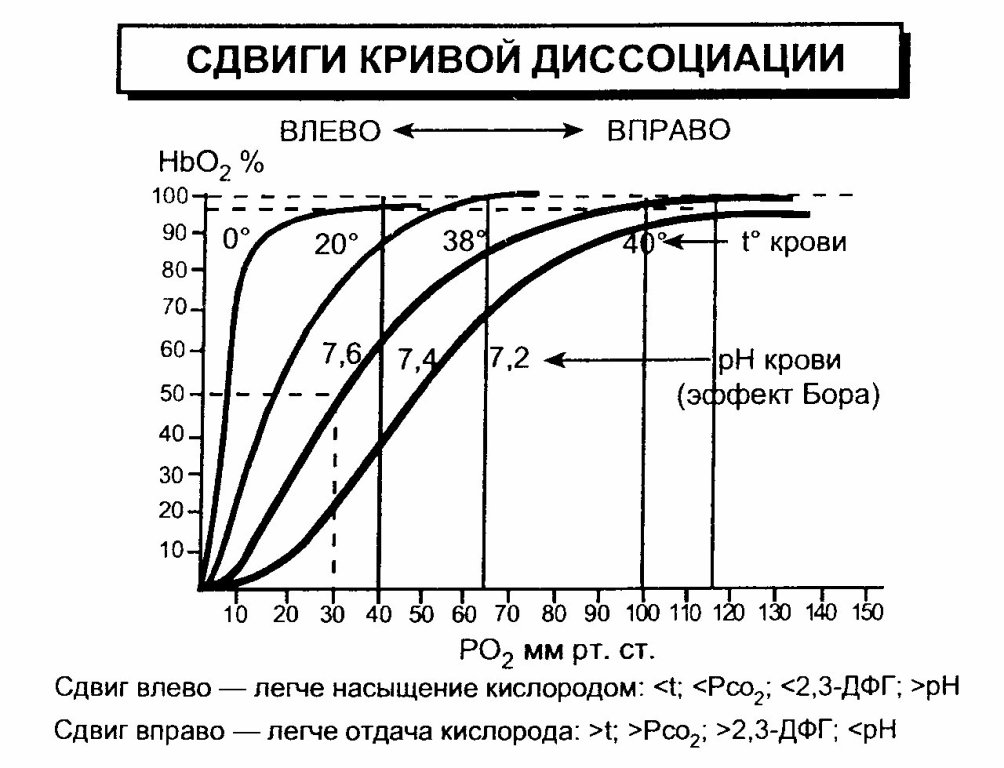
Оксигенированной артериальной крови соответствует плато кривой диссоциации, а десатурированной крови в тканях — круто снижающаяся ее часть. Пологий подъем кривой в верхнем ее участке (зона высокого напряжения О2) свидетельствует, что достаточно полное насыщение гемоглобина артериальной крови кислородом обеспечивается даже при уменьшении напряжения 02 до 70 мм рт.ст. Понижение напряжения О2 со 100 на 15-20 мм рт. ст. практически не отражается на насыщении гемоглобина кислородом (НЬО; снижается при этом на 2-3%). При более низких значениях напряжения О2 оксигемоглобин диссоциирует значительно легче (зона крутого падения кривой). Так, при снижении напряжения 02 с 60 до 40 мм рт. ст. насыщение гемоглобина кислородом уменьшается приблизительно на 15%. Положение кривой диссоциации оксигемоглобина количественно принято выражать парциальным напряжением кислорода, при котором насыщение гемоглобина составляет 50%. Нормальная величина Р50 при температуре 37°С и рН 7.40 — около 26.5 мм рт. ст.. Кривая диссоциации оксигемоглобина при определенных условиях может смещаться в ту или иную сторону, сохраняя S-образную форму, под влиянием изменения: 1. рН, 2. напряжения СО, 3. температуры тела, 4. содержания в эритроцитах 2,3-дифосфоглицерата (2,3-ДФГ), от которых зависит способность гемоглобина связывать кислород. В работающих мышцах в результате интенсивного метаболизма повышается образование СО2 и молочной кислоты, а также возрастает теплопродукция. Все эти факторы понижают сродство гемоглобина к кислороду. Кривая диссоциации при этом сдвигается вправо, что приводит к более легкому освобождению кислорода из оксигемоглобина, и возможность потребления тканями кислорода увеличивается. При уменьшении температуры, 2,3-ДФГ, снижении напряжения СО2 и увеличении рН кривая диссоциации сдвигается влево, сродство гемоглобина к кислороду возрастает, в результате чего доставка кислорода к тканям уменьшается.
6. Транспорт углекислого газа кровью. Углекислый газ переносится к легким в форме бикарбонатов и в состоянии химической связи с гемоглобином (карбогемоглобин).
Углекислый газ является продуктом метаболизма клеток тканей и поэтому переносится кровью от тканей к легким. Углекислый газ выполняет жизненно важную роль в поддержании во внутренних средах организма уровня рН механизмами кислотно-основного равновесия. Поэтому транспорт углекислого газа кровью тесно взаимосвязан с этими механизмами. В плазме крови небольшое количество углекислого газа находится в растворенном состоянии; при РС02= 40 мм рт. ст. переносится 2,5 мл/100 мл крови углекислого газа, или 5 %. Количество растворенного в плазме углекислого газа в линейной зависимости возрастает от уровня РС02. В плазме крови углекислый газ реагирует с водой с образованием Н+ и HCO3. Увеличение напряжения углекислого газа в плазме крови вызывает уменьшение величины ее рН. Напряжение углекислого газа в плазме крови может быть изменено функцией внешнего дыхания, а количество ионов водорода или рН — буферными системами крови и HCO3, например путем их выведения через почки с мочой. Величина рН плазмы крови зависит от соотношения концентрации растворенного в ней углекислого газа и ионов бикарбоната. В виде бикарбоната плазмой крови, т. е. в химически связанном состоянии, переносится основное количество углекислого газа — порядка 45 мл/100 мл крови, или до 90 %. Эритроцитами в виде карбаминового соединения с белками гемоглобина транспортируется примерно 2,5 мл/100 мл крови углекислого газа, или 5 %. Транспорт углекислого газа кровью от тканей к легким в указанных формах не связан с явлением насыщения, как при транспорте кислорода, т. е. чем больше образуется углекислого газа, тем большее его количество транспортируется от тканей к легким. Однако между парциальным давлением углекислого газа в крови и количеством переносимого кровью углекислого газа имеется криволинейная зависимость: кривая диссоциации углекислого газа. Роль эритроцитов в транспорте углекислого газа. Эффект Холдена. В крови капилляров тканей организма напряжение углекислого газа составляет 5,3 кПа (40 мм рт. ст.), а в самих тканях — 8,0—10,7 кПа (60—80 мм рт. ст.). В результате С02 диффундирует из тканей в плазму крови, а из нее — в эритроциты по градиенту парциального давления С02. В эритроцитах С02 образует с водой угольную кислоту, которая диссоциирует на Н+ и HCO3. (С02 + Н20 = Н2СО3 = Н+ + HCO3). Эта реакция протекает быстро, поскольку С02 + Н20 = Н2СО3 катализируется ферментом карбоангидразой мембраны эритроцитов, которая содержится в них в высокой концентрации. В эритроцитах диссоциация углекислого газа продолжается постоянно по мере образования продуктов этой реакции, поскольку молекулы гемоглобина действуют как буферное соединение, связывая положительно заряженные ионы водорода. В эритроцитах по мере освобождения кислорода из гемоглобина его молекулы будут связываться с ионами водорода (С02 + Н20 = Н2С03 = = Н+ + HCO3), образуя соединение (Нb-Н+). В целом это называется эффектом Холдена, который приводит к сдвигу кривой диссоциации оксигемоглобина вправо по оси х, что снижает сродство гемоглобина к кислороду и способствует более интенсивному освобождению его из эритроцитов в ткани. При этом в составе соединения НЬ-Н+ транспортируется примерно 200 мл С02 в одном литре крови от тканей к легким. Диссоциация углекислого газа в эритроцитах может быть лимитирована только буферной емкостью молекул гемоглобина. Образующиеся внутри эритроцитов в результате диссоциации С02 ионы НСОз с помощью специального белка-переносчика мембраны эритроцитов выводятся из эритроцитов в плазму, а на их место из плазмы крови закачиваются ионы Сl- (феномен «хлорного» сдвига). Основная роль реакции С02 внутри эритроцитов заключается в обмене ионами Сl- и НСОз между плазмой и внутренней средой эритроцитов. В результате этого обмена продукты диссоциации углекислого газа Н+ и НСОз будут транспортироваться внутри эритроцитов в виде соединения (Нb-Н+), а плазмой крови — в виде бикарбонатов. Эритроциты участвуют в транспорте углекислого газа от тканей к легким, поскольку С02 образует прямую комбинацию с — NН2-группами белковых субъединиц гемоглобина: С02 + Нb -> НbС02 или карбаминовое соединение. Транспорт кровью С02 в виде карбаминового соединения и ионов водорода гемоглобином зависит от свойств молекул последнего; обе реакции обусловлены величиной парциального давления кислорода в плазме крови на основе эффекта Холдена. В количественном отношении транспорт углекислого газа в растворенной форме и в форме карбаминового соединения является незначительным, по сравнению с его переносом С02 кровью в виде бикарбонатов. Однако при газообмене С02 в легких между кровью и альвеолярным воздухом эти две формы приобретают основное значение. Когда венозная кровь возвращается от тканей к легким, С02 диффундирует из крови в альвеолы и РС02 в крови снижается с 46 мм рт. ст. (венозная кровь) до 40 мм рт.ст. (артериальная кровь). При этом в величине общего количества С02 (6 мл/100 мл крови), диффундирующего из крови в альвеолы, доля растворенной формы С02 и карбаминовых соединений становится более значительной относительно бикарбонатной. Так, доля растворенной формы составляет 0,6 мл/100 мл крови, или 10 %, карбаминовых соединений — 1,8 мл/100 мл крови, или 30%, а бикарбонатов — 3,6 мл/100 мл крови, или 60 %. В эритроцитах капилляров легких по мере насыщения молекул гемоглобина кислородом начинают освобождаться ионы водорода, диссоциировать карбаминовые соединения и НСОз вновь превращается в С02 (Н+ + НСОз = = Н2С03 = С02 +Н20), который путем диффузии выводится через легкие по градиенту его парциальных давлений между венозной кровью и альвеолярным пространством. Таким образом, гемоглобин эритроцитов играет основную роль в транспорте кислорода от легких к тканям, и углекислого газа в обратном направлении, поскольку способен связываться с 02 и Н+. В состоянии покоя через легкие из организма человека за минуту удаляется примерно 300 мл С02: 6 мл/100 мл крови х 5000 мл/мин минутного объема кровообращения.
7. Регуляция дыхания. Дыхательный центр, его отделы. Автоматия дыхательного центра. Хорошо известно, что внешнее дыхание постоянно изменяется в различных условиях жизнедеятельности организма. Дыхательная потребность. Деятельность функциональной системы дыхания всегда подчинена удовлетворению дыхательной потребности организма, которая в значительной степени определяется тканевым метаболизмом. Так, при мышечной работе по сравнению с покоем возрастает потребность в кислороде и удалении двуокиси углерода. Для компенсации повышенной дыхательной потребности увеличивается интенсивность легочной вентиляции, что выражается в увеличении частоты и глубины дыхания. Роль двуокиси углерода. Эксперименты на животных показали, что избыток двуокиси углерода в воздухе и крови (гиперкапния) стимулирует легочную вентиляцию за счет учащения и углубления дыхания, создавая условия для удаления из организма ее избытка. Напротив, снижение парциального давления двуокиси углерода в крови (гипокапния) вызывает уменьшение легочной вентиляции вплоть до полной остановки дыхания (апноэ). Это явление наблюдается после произвольной или искусственной гипервентиляции, во время которой из организма в избытке удаляется двуокись углерода. В результате сразу же после интенсивной гипервентиляции возникает остановка дыхания — постгипервентиляционное апноэ. Роль кислорода. Недостаток кислорода в атмосфере, снижение его парциального давления при дыхании на большой высоте в условиях разреженной атмосферы (гипоксия) также стимулируют дыхание, вызывая увеличение глубины и особенно частоты дыхания. В результате гипервентиляции недостаток кислорода частично компенсируется. Избыток кислорода в атмосфере (гипероксия), наоборот, снижает объем легочной вентиляции. Во всех случаях вентиляция изменяется в направлении, способствующем восстановлению измененного газового состояния организма. Процесс, называемый регуляцией дыхания, заключается в стабилизации дыхательных показателей у человека. Под главным дыхательным центром понимают совокупность нейронов специфических дыхательных ядер продолговатого мозга. Дыхательный центр управляет двумя основными функциями; двигательной, которая проявляется в виде сокращения дыхательных мышц, и гомеостатической, связанной с поддержанием постоянства внутренней среды организма при сдвигах в ней содержания 02 и С02 Двигательная, или моторная, функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна. Благодаря этой функции осуществляется интеграция дыхания с другими функциями. Под паттерном дыхания следует иметь в виду длительность вдоха и выдоха, величину дыхательного объема, минутного объема дыхания. Гомеостатическая функция дыхательного центра поддерживает стабильные величины дыхательных газов в крови и внеклеточной жидкости мозга, адаптирует дыхательную функцию к условиям измененной газовой среды и другим факторам среды обитания. Дата добавления: 2015-12-15 | Просмотры: 1337 | Нарушение авторских прав |