|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ГАМЕТОГЕНЕЗ И МЕЙОЗПроцесс развития половых клеток носит название гаметогене-за. У самцов этот процесс называется спермиогенезом, а у самок — овогенезом (рис. 6). Половые клетки в развитии последовательно проходят следующие стадии: размножения, роста, созревания и формирования. В стадии размножения клетки интенсивно делятся митотическим путем. В стадии роста клетки накапливают питательные вещества, особенно при овогенезе. Наиболее ответственный момент с точки зрения генетики при образовании половых клеток — мейоз — процесс редукционного и эквационного деления ооцитов и сперматоцитов, в результате которого образуются половые клетки с гаплоидным набором хромосом. Рассмотрим наиболее важные моменты поведения хОомосом в мейозе. В этом процессе выделяют две стадии деления (см. схему): 1) редукционную и 2) эквационную. Схема мейоза
редукционное деление профаза I — лептонема зигонема пахинема диплонема диакинез метафаза I анафаза I телофаза I Образование из одной материнской RXB с диплоидным набором хромо- ДЧ«дочерних с гаплоидным набо- удвоенных хромосом
ЗИГОТА Рис. 6. Сравнение процессов сперматогенеза и овогенеза у животных с гаплоидным числом хромосом, равным 2 (по К. Вилли и В. Детье, 1975) Непосредственно перед мейозом клетки половых желез находятся в интерфазе. Редукционное деление начинается с профазы I, которая, как видно из схемы, подразделяется на пять фаз. На первой стадии профазы I — лептонемы хромосомы деспирализо-ваны, они в 2—5 раз длиннее метафазных. Под электронным микроскопом можно видеть, что они состоят из двух хроматид, соединенных центромерой. На следующей стадии — зигонемы наблюдаются притяжение и слияние (конъюгация) гомологичных хромосом. Каждая пара конъюгирующих хромосом образует бивалент, а по числу хроматид — тетраду. На этой стадии происходит образование синаптонемного комплекса (СК), входящего в состав бивалента. Нарушение формирования СК между гомологичными хромосомами наблюдают у гетерозиготных носителей хромосомных аберраций. Далее, на стадии пахинемы, происходят утолщение и укорочение хромосом, так что сестринские хроматиды становятся хорошо различимыми; на отдельных из них можно видеть и ядрышки. Следующая фаза — диплонема характеризуется тем, что конъ-югярующие хромосомы начинают отталкиваться и постепенно расходятся от центромеры к концам. При этом образуются характерные фигуры, напоминающие греческую букву «хи» (х) и получившие вследствие этого название хиазмы. В точках соприкосновения гомологичных хромосом возникают разрывы. Они могут быть одинарными, двойными и более сложными. В результате разрывов образуются фрагменты хроматид, которые затем могут воссоединяться на другой хромосоме, изменяя тем самым комбинацию генетического материала в клетке. Обмен участками между гомологичными хромосомами получил название кроссинговера. На последней стадии профазы I — диакинезе происходит резкое укорочение хромосом, так что к концу этой стадии хроматиды остаются связанными только на концевых участках. Этим и заканчивается профаза I. Необходимо отметить, что при более детализированном изучении мейоза в профазе выделяют и другие промежуточные стадии, например пролептонему, диктионему и т. д. На стадии метафазы I биваленты располагаются в плоскости экватора центромерами к противоположным полюсам. Силы отталкивания здесь увеличиваются. В анафазе I начинается расхождение гомологичных хромосом к противоположным полюсам, которое носит случайный характер. Каждая из пар гомологичных хромосом имеет одинаковую вероятность распределения в одну из двух дочерних клеток. В телофазе I хромосомы достигают полюсов клетки. Затем восстанавливаются ядерная оболочка и ядрышко, хромосомы декон-денсируются. В конце телофазы делится цитоплазма {цитокинез) и образуются две дочерние клетки с гаплоидным набором хромосом. Отличительной особенностью первой стадии мейоза является то, что в период анафазы сами хромосомы не делятся на хроматиды, как при митозе, а лишь расходятся гомологичные пары хромосом к разным полюсам клетки и формируются две дочерние клетки с редуцированным наполовину набором хромосом, состоящим, однако, из двух хроматид. Между первой и второй стадиями мейоза имеется непродолжительный период покоя — интеркинез, во время которого не Происходит репродукции хромосом. Эквационное, или уравнительное, деление ^илогично митозу, где клетки последовательно проходят четыре Фазы: профазу II, метафазу II, анафазу II, телофазу П. На стадии °*шфазы II хромосомы разделяются на две хроматиды, которые затем с помощью нитей веретена расходятся к противоположным полюсам. На стадии телофазы II заканчивается формирование
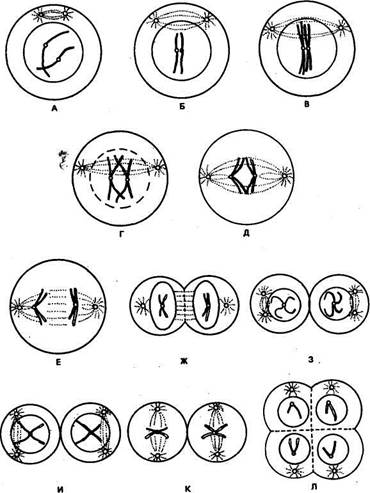
Рис. 7, Схем» последомтелных стадий мейоза (по К. Свенсону и П. Уэбстеру, 1980): А — лептонема, предшествующая конъюгации хромосом; Б — начало конъюгации на стадии зигонемы; В— пахинема; Г— диплонема; Д— метафаза I; E— анафаза I; Ж~- телофаза I; 3 — интерфаза между двумя делениями мейоза; И— профаза II; К— метафаза II; Л— тело-фаза II Таким образом, в результате двух меиотических делений из одной клетки с диплоидным набором хромосом образуются четыре клетки с гаплоидным набором хромосом и в 2 раза меньшим, чем в соматических клетках, содержанием ДНК. Вероятностный характер распределения материнских и отцовских гомологичных хромосом в разные клетки позволяет создать новые комбинации негомологичных хромосом в яйцеклетках и сперми-ях, чем достигается огромное число новых сочетаний наследственной информации. Новые сочетания генетической информации возникают вследствие кроссинговера. Каждая из хромосом в метафазе I содержит участки, происходящие от отцовских и материнских хромосом. Рекомбинации хромосом при кроссинговере и вероятностное распределение их по клеткам — причины наследственной изменчивости организма. Мейоз, оплодотворение и митоз обеспечивают поддержание постоянства числа хромосом в смежных поколениях видов. В этом их биологическое значение. Патология мейоза. Основная патология мейоза — нерасхождение хромосом- Оно может быть первичным, вторичным и третичным. Первичное нерасхождение возникает у особей с нормальным кариотипом. При этом на стадии анафазы I нарушается разделение бивалентов и обе хромосомы из пары гомологов переходят в одну клетку, что приводит к избытку хромосом в данной клетке (и + 1) и недостатку в другой (и—1). Вторичное нерасхождение возникает в гаметах у особей с избытком (трисо-мией) одной хромосомы в кариотипе. В результате этого в процессе мейоза образуются и биваленты, и униваленты. Третичные нерасхождения наблюдают у особей, имеющих структурные перестройки хромосом, например транслокации. Нерасхождение хромосом отрицательно влияет на жизнеспособность организма животных. Подробно об этом будет изложено в последующих главах. Отличительная особенность мейоза у самок — образование в первом и втором меиотических делениях так назьгааемых полярных телец, которые впоследствии дегенерируют и в размножении не участвуют. Неравные деления в овогенезе обеспечивают яйцеклетке необходимое количество цитоплазмы и запасного желтка, чтобы она могла выжить после оплодотворения. В отличие от спермиогенеза, который у самцов происходит как во внутриутробный (пренатальный) период, так и после рождения (постна-тальный период), у самок яйцеклетка после рождения не образуется. Однако к концу пренатального периода у самок накапливается огромное количество овоцитов (у коров, например, десятки тысяч), а созревают и дают начало развитию потомства лишь немногие из них. Этот резерв генетического материала, особенно у малоплодных животных, таких, как крупный рогатый скот, в настоящее время начинают использовать для искусственного стимулирования созревания многих яйцеклеток, последующего их оплодотворения и пересадки (трансплантации) специально подготовленным коровам-реципиентам. Таким образом от одной ценной коровы в год можно получить не одного, а несколько телят. Оплодотворение наступает после слияния гаплоидных сперматозоида и яйцеклетки и образования диплоидной клетки — зиготы, дающей начало развитию эмбриона. При делении клеток эмбриона, содержащих хромосомы матери и отца, генетическая информация поступает во все клетки нового организма. Морфологические и функциональные различия хромосомных наборов — основные причины эмбриональной смертности после оплодотворения при скрещивании разных видов или бесплодия гибридов. Так, при скрещивании зайца и кролика не происходит имплантации и развития оплодотворенной яйцеклетки. Эмбриональная смертность наступает при скрещивании козы и овцы. Мужские гибвиды осла (2 л = 62) и лошади (2л = 64), как известно, не дф.т потомства, они стерильные, или бесплодные. Стерильность гибридов-самцов наблюдается при скрещивании европейского крупного рогатого скота с бизоном, зубром, яком, а также гауром и гаялом. Это связано с тем, что у гибридов нарушаются процессы спермиогенеза. Однако при скрещивании европейского крупного рогатого скота с азиатским горбатым зебу, дикого кабана (2л = 36) и домашней свиньи (2л = 38) потомство рождается плодовитое. В результате сложных вариантов скрещиваний в последнее время удалось получить плодовитых гибридов крупного рогатого скота с зуС^ом, бизоном, яком. Гибриды отличаются такими ценными качествами, как крепкое телосложение, высокая жизнеспособность, хорошие мясные признаки. Контрольные вопросы. 1. Каковы морфологическое строение и химический состав хромосом? 2. Что такое кариотип и каковы его особенности у разных видов животных? 3. Что такое митоз и каково его биологическое значение? 4. Что такое гаметогенез и каковы его особенности у самцов и самок? 5. Что вы знаете о мейозе и его биологическом значении? 6. Каковы основные формы патологий митоза и мейоза? Глава 3 Дата добавления: 2015-12-16 | Просмотры: 1278 | Нарушение авторских прав |



 еще двух клеток. В результате после двух последовательных стадий мейоза из каждой клетки образуются четыре новые с гаплоидным набором хромосом. Для более наглядного представления всех этих событий можно воспользоваться схемой мейоза, приведенной на рисунке 7.
еще двух клеток. В результате после двух последовательных стадий мейоза из каждой клетки образуются четыре новые с гаплоидным набором хромосом. Для более наглядного представления всех этих событий можно воспользоваться схемой мейоза, приведенной на рисунке 7.