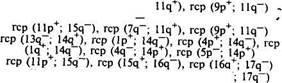|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Хряков-носителей реципрокных транслокаций и эмбриональнойСмертности у их потомства
несбалансированных кариотипов, %
13 iZq- Рис. 60. Транслокация между 13-й и 14-й хромосомами свиньи (по Хагельдорну и др.) 51. Частота вовлечения хромосом свиней в спонтанные реципрокные транслокации
Р i 6q), rcpjlp; 14qx), гср
5q), гср (lq_; 17q+), rep (lq+
7q), rep (lp-; 8ql
rep (3p+- 7q~) rep (4q; 14p), гср (4q+; 13q), гср (4q; 15p+)
rep (5q; 8q+), rep (5p~; 14p+) rep (6p; 15q), rep (6p+; 14q~), гср (lp-; 6q+) гср (7q; llq), rep (7pJ; 13q~), rep (3p; 7~), r dq; 7q)
rep (lp-; 8q+) rep (9p+; llq-) гср (llp+; 15q~) rcp(13q-; 14p+) гср (4q"; 14p+) гср (9р+; llq-) шведской йоркширской породы, гетерозиготного по транслокации (13q~; 14q+), на стадии диакинез — метафаза I наблюдали 17 бивалентов, по одному квадриваленту. У свинок наблюдали хромосомы с кольцевой конфигурацией, что характеризует реци-прокную транслокацию. У хрячков кольцевую конфигурацию наблюдали примерно в 20 % клеток на стадии диакинез — метафаза I. Образование квадривалентов приводит к формированию несбалансированных гамет на стадии метафазы И, что является причиной нарушений процессов эмбриогенеза после оплодотворения. Получены четкие доказательства участия гамет с несбалансированным кариотипом в оплодотворении, что свидетельствует о неэффективности или отсутствии презиготического отбора (элиминации гамет с несбалансированным кариотипом до оплодотворения). Исследовали предимплантационные эмбрионы, полученные после случки гетерозиготных по реципрокной (13q~~; 14q+) транслокации хряков с 10 нормальными свинками. В 36 карио-типированных эмбрионах 14 имели нормальный сбалансированный набор хромосом, 11 — сбалансированный, но гетерозиготный по транслокации, 11— несбалансированный. В последнем случае у эмбрионов кроме структурной перестройки — транслокации были выражены и изменения числа хромосом в кариоти-пе — трисомия или моносомия как следствие нарушения их расхождения в мейозе. Во втором варианте исследования случали хряка с нормальным набором хромосом с 10 гетерозиготными свинками. Цитбгенетический анализ проведен у 33 эмбрионов. Из них 10 оказались с нормально сбалансированным кариотипом, 14 гетерозиготных сбалансированных, 9 имели несбалансированный набор хромосом. В третьем варианте хряка случали с тремя нормальными свинками. В результате анализа выявлено 17 нормальных сбалансированных и 12 сбалансированных, но гетерозиготных эмбрионов. В четвертом варианте после случки нормального хряка и четырех гетерозиготных свинок изучено 17 эмбрионов, из которых 5 имели нормальный кариотип, 10 оказались гетерозиготными сбалансированными и 2 дегенерированными. В последнем варианте гетерозиготного хряка случали с гетерозиготной свинкой. Из четырех проанализированных эмбрионов 2 имели сбалансированный с гетерозиготной формой транслокации кариотип и 2 эмбриона были дегенерированные. Эмбриональная смертность, вычисленная по числу желтых тел и живых имплантированных эмбрионов, у гетерозиготных свинок, слученных с нормальными хряками, а также у нормальных и гетерозиготных свинок, слученных с гетерозиготными хряками, составила соответственно 72,3; 68,0; 85,5 %. Таким образом, у хряков и свинок — носителей транслокации хромосом наблюдаются нарушения процессов мейоза и гаметогенеза. Такие спер-мии и яйцеклетки, участвуя в оплодотворении, дают нежизнеспособных эмбрионов, что выражается в резком снижении количества поросят в пометах. Рещрпхжную транслокацию Т (llp+; 15q~) в гетерозиготном состоянии обнаружил Кинг у двух хряков — отца и сына — со сниженной на 56 и 34 % оплодотворяющей способностью. Эта транслокация в сбалансированной форме обнаружена у 41 % живых потомков сына. Несбалансированный кариотип выявлен у 11 % эмбрионов, изученных в период от 10-го до 88-го дня беременности. Эти эмбрионы, несомненно, погибают. 100%-ную эмбриональную смертность наблюдали Бойтерс и др. в потомстве при использовании хряка — мозаика по реципрокной транслокации между 6-й и 15-й хромосомами. Кариотип этого животного был 38, XY/38, XY+ (6q+; 15q). Высокую степень смертности, включая мертворожденных поросят и потомков с врожденными уродствами, зарегистрировали Попеску и Легатт у хряка, в кариотипе которого обнаружено 24 % клеток со структурными перестройками. Подтверждением того положения, что снижение плодовитости у свиней может быть связано с нарушением генома, является сообщение Фогта и др. Два хряка-брата, дающих потомство с 50%-ным уменьшенным размером помета, в сравнении с другими хряками и теми же свиноматками оказались миксоплоида-ми -37, XY-18/38, XY/39, XY, +18 и 37, XY, - 18/38, XY. При анализе кариотипа их потомков у двух свиней-дочерей с пониженной плодовитостью был обнаружен аномальный набор хромосом. Норби с соавт. при анализе бластоцист у свиней в 10' % случаев из 38 обнаружили количественные и структурные аномалии, в их числе 4 триплоида, 3 тетраплоида, 1 диплоид/триплоид и <здна деления. При изучении 13 хряков с пониженной плодовитостью и 100%-ной эмбриональной смертностью потомства Бойтерс установил, что основная причина нарушения эмбриоге- неза — аномалии кариотипа: транслокации, палочковидные хромосомы, которые наблюдались в 25 % из 40 % эмбрионов, созревших до 25 дней. В исследованиях М. Л. Кочневой и Н. А. Осиповой (1995) показано увеличение соматической хромосомной нестабильности у поросят с врожденными аномалиями и у свиноматок с низкой плодовитостью. Цитогенетический анализ у свиней дал возможность изучить связь нарушений плодовитости с аномалиями в системе половых хромосом. Так, на метафазных препаратах поросят-интерсексов был обнаружен химеризм XX/XY и XX/XXY. Такая же аномалия хромосом в лимфоцитах найдена у плодовитой свиноматки (среднее число поросят в один помет — 14), у которой, по данным Кристенсена, 24 % клеток имели половые хромосомы XY, остальные содержали ХХ-хромосомы. У свиней обнаружена аномалия кариотипа, сходная с синдромом Клайнфельтера у человека и ранее описанная на других видах (2n-39,XXY). Для характеристики спермиогенеза у этого животного исследовали суспензию эпидидимальных клеток и семенники. В содержимом эпидидимуса спермин обнаружены не были. Гистологический анализ, проведенный Ханкоком, показал отсутствие спермато-генного эпителия. У свиней зарегистрировано рождение поросят с одной половой хромосомой 37, ХО, что соответствует синдрому Тернера у человека. Значительное количество цитогенетических исследований проведено у свиней истинных и псевдогермафродитов. При этом в разных тканях (кровь, кожа, костный мозг) обнаружены различные хромосомные наборы, в том числе нормальные, химеризм по половым хромосомам, иногда в сочетании с трисомией или моносомией - 38, ХХ/39, XXY; 37, ХО/38, ХХ/38, XY, отдельно трисомия 39, XXY. Представляют интерес сообщения Михельмана и соавт. о различиях частот структурных и морфологических изменений хромосом в группах племенных хряков и свиноматок в сравнении с откормочными выбракованными животными. В первой группе структурные изменения обнаружены в 17,9 %, во второй — в 13,7 и в третьей — в 20,7 % метафаз. Морфологические изменения хромосом наблюдали соответственно в 0,58; 0,61 и 0,72 % метафаз. Аналогичные результаты получены и у крупного рогатого скота. Это указывает на то, что снижение плодовитости и болезни как основные причины выбраковки животных связаны не только с действием факторов внешней среды. Первичная причина нарушений функций организма может быть связана с изменениями в генотипах животных. Овцы. Нормальный кариотип овец состоит из 54 хромосом, что впервые указано советским генетиком Живаго (1931). У
овец, как и у других видов животных, обнаружены различные виды аберраций хромосом. Так, в Новой Зеландии, в стране с развитым овцеводством, наблюдали три типа<робертсоновской транслокации — между 5-й и 26-й, 8-й и 11-й, 7-й и 25-й хромосомами, получившими название Массей I, Массей II и Массей III. Однако, как показали Бруер и Чепман, плодовитость у животных была нормальной. Это объясняется естественной выбраковкой не сбалансированных по числу хромосом гаплоидных клеток еще до вступления их в оплодотворение. У овец описаны также отдельные варианты реципрокных транслокаций, которые сопровождались пониженной плодовитостью животных. В Институте экспериментальной биологии Казахстана установлено, что у каракульских баранов в очень молодом и старом возрастах хромосомные аберрации в генеративной ткани встречаются значительно чаще, чем в среднем возрасте. Хромосомные аберрации были обнаружены у некоторых мертворожденных с врожденными аномалиями ягнят. Лошади. Нормальный кариотип домашней лошади состоит из 64 хромосом. Точное число хромосом этого вида животных установили Сасаки и Макино только в 1962 г. Количество цитогене-тических исследований у лошадей пока еще невелико, поэтому установить истинную частоту аберраций хромосом не представляется возможным. Однако Виллер и Визнер (1981) считают, что по крайней мере 5—10 % всех зигот у лошадей, как и у других видов млекопитающих животных, могут содержать хромосомные аномалии. 90 % этих зигот отмирают на первой стадии беременности. Исследования кариотипа лошадей показывают, что наиболее часто у этого вида встречаются аномалии в системе половых хромосом, которые вызывают бесплодие у животных (табл. 53). Дата добавления: 2015-12-16 | Просмотры: 902 | Нарушение авторских прав |
 Частота
Частота


 Число транслокаиий
Число транслокаиий