|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
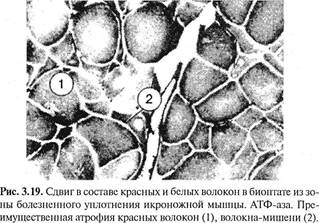
ВЕРТЕБРОНЕВРОЛОГИЧЕСКОГО 5 страницаВ участках измененных мышечных волокон соединительная ткань была разрыхлена, отечна, иногда с увеличением количества клеточных элементов и коллагеновых волокон, местами с переходом в фиброз и склероз. В таких участках мышечные волокна были раздвинуты. Соединительная ткань, богатая клеточными элементами, гистиоцитами, фибробластами и лимфоидными элементами, иногда имела рубцовый вид. В некоторых сосудах эндотелий был набухший, иногда пролиферирующий. Отмечалось утолщение стенок сосудов. В мышечных волокнах этих участков обнаруживались выраженные явления атрофии и дистрофии, в некоторых из них встречались многочисленные ядра и клетки (рис. 3.18). Подобные изменения были описаны затем и H.Tichy, K.Seidel (1969). Таким образом, вторая (органическая) стадия мышечно-болевого синдрома связана с различными дистрофическими изменениями: зернистый и глыбчатый распад, очаговый миолиз, дисковидный распад мышечных волокон, вакуольная дистрофия и пр., с переходом в фиброз и склероз. Биопсийный материал, взятый из болезненных мышечных уплотнений икроножной мышцы у 23 больных вертеброгенной люмбоишиальгией, также показал, что вслед за серозным пропитыванием наступают продуктивно-пролиферативные изменения. Обнаруживались группы одиночных атрофированных мышечных волокон, волокна-мишени, фиброз эндомизия, а в поздних стадиях — более грубый фиброз (Попелянский Я.Ю., Хаби-ровФ.А., 1993). Укороченная мышца теряет уровень диффе-ренцировки, происходит сдвиг в составе «медленных» и «быстрых» волокон (рис. 3.19). J.Travell и D.Simons (1983) считают укорочение специфическим фактором активации курковых зон мышцы: длительное укорочение, например во сне, или внезапное, например при расслаблении укороченного агониста хлорэтилом. Огра- Глава ILL Методики вертеброневрологического обследования
ничение функции характерно как для активного, так и для латентного пунктов, спонтанная же боль — только для первого. Белки мышечно-сухожильной ткани и болезненных мышечных узелков — зон нейроостеофиброза становятся ауто-антигенами (Tichy Н., Seidel К., 1985; Попелянский Я.Ю., Ве-селовская О.И., 1985). Перед тем как указать на малотравматические современные методики оценки состояния мышц при болевых синдромах, соответствующие данные целесообразно сопоставить с результатами экспериментальных исследований на животных. С.С.Вайль (1967) вызвал перенапряжение мышц, вынуждая крыс плавать до изнеможения. При окраске мышц этих животных гематоксилин-эозином в поляризованном свете мышечные волокна определялись слегка извилистыми, саркоплазма образовывала горбовидные выбухания. В соответствующих участках анизотропные диски сливались, образуя сплошные двояколучепреломляющие фрагменты, что, по мнению автора, соответствует выраженным контрактурам. Подобные изменения находил наш сотрудник В.П.Веселов-ский после такого же типа перенапряжения мышц у подопытных животных. Другой группе крыс в латеральную головку икроножной мышцы вводилось 0,5 мл 10% раствора хлористого кальция. На втором месяце опыта в толще икроножной мышцы, чаще в месте начала обеих головок, выявлялась болезненность — животные реагировали на их пальпацию более бурно, чем на пальпацию других зон. Начиная с четвертого месяца, в медиальной головке мышцы обнаруживались постоянные участки уплотнения, и на той же конечности определялась кожная гипотермия. Здесь же на электромиограмме регистрировалось удлинение периода «успокоения» в ответ на погружение игольчатого электрода в мышцу. Выявлялась также нейрогуморальная декомпенсация в системе ацетилхолин-холинэстераза (гиперацетилхолинемия при снижении активности холинэстеразы), увеличивалась активность окислительно-восстановительных ферментов, лактатдегидрогеназы и сукцинатдегидрогеназы, снижалась активность ДПН-диа-форазы (коэнзимы I и II, динуклеатиды фосфатные). Нарушения в указанных системах находились в прямой зависимости от выраженности уплотнений в мышцах. В свете изложенного представляют интерес результаты исследований наших учеников по изучению обсуждаемой патологии мышц. В альгической фазе, когда объективные симптомы ограничиваются почти исключительно местными изменениями тонуса мышцы, интересные показатели нередко дает игольчатая электромиография. Обнаруживаются усиление и удлинение периода биоэлектрической активности погружения и усиление активности покоя. Появляется залпообразная активность покоя в ограниченных участках гипертонуса. В этот же период улавливаются и сдвиги тканевого клиренса (процента снижения радиоактивности в минуту) при введении изотопа в пораженную мышцу (Аляби-на Н.Е., Заславский Е.С., Ходич Т.Г. и соавш., 1973). Что касается нейродистрофических влияний на мышцу и соединительную ткань, то на первых порах предполагалось, что они адресуются к сосудистому аппарату и реализуются через вазомоторику (Сеченов И.М., 1866; Бехтерев М., 1925; Brunning L., 1952). Одновременно на разных этапах
изучения нейротрофики накапливались факты в пользу непосредственных нервно-трофических влияний. В лаборатории С.П.Боткина его сотрудники С.В.Левашов (1880), а затем И.П.Павлов (1882,1883) наблюдали усиление функциональной активности стенки сосудов задней лапы и сердечной мышцы собаки при раздражении определенных нервов. Это в последующем (1892) позволило И.П.Павлову поддержать представление S.Magandie (1834), Samuel'a (1860) о наличии специальных трофических нервов. Эту рефлекторную функцию стали приписывать всей вегетативной или только симпатической (Орбели Л.А., 1938), или соматической нервной системе1. В последнем случае одни авторы отдавали преимущество чувствительной (Wybum-Mason R., 1950; Грацианская A.M., 1956; Григорьева Т.А., 1959; Лебединский А.В., Нахильницкая З.Н., 1964), другие — двигательной иннервации (Gutman Е. et al, 1956, 1962; Гник П., 1958; Hollos G. et al., 1960; Felix W., 1962). Л.А.Орбели (1938) говорил о тономоторной функции симпатикуса, создающей почву для действий соматических тономоторных нервов. М.Р.Могендович отождествляет эту концепцию с представлением R.Granit (1957) о у-эфферентах. В настоящее время большинство авторов признает, что трофическая функция осуществляется всей нервной системой, а нарушение трофики может возникнуть при повреждении любого ее отдела (Сперанский А.Д., 1936, 1952; Баяндуров Б. И., 1949; Коштоянц Х.С, 1951; Боровский М.Л., 1952, 1957; Зай-ко Н.Н., 1954; Бентелев A.M., Надежкин Л.В., 1959; Волкова О.В., 1970; Швалев В.К, 1971). Прямые трофические влияния дополняются влиянием симпатикуса на сосуды, питающие ткань через адренергические волокна (Говырин В.Н., 1967). Эти общие теоретические представления о нейродистро-фии позволили глубже оценить новые факты, полученные благодаря внедрению внутриклеточной регистрации электрических потенциалов, гистохимического и электронно-микроскопического методов, осмыслить изменения в клетках на молекулярном и субклеточном уровнях. Общие синтетического характера концепции не препятствовали выяснению интимных механизмов реализации трофических функций. В частности, продолжается раздельное изучение возбуждающего действия нерва, т.е. приводящего к мышечному сокращению, и трофического. 1 Важным этапом изучения вегетативной нервной системы был классический эксперимент Гинецинского-Орбели на нервно-мышечном препарате: раздражение симпатических связей предотвращало трофическое истощение мышцы. Экспериментальное оправдание концепции Л.А.Орбели (1939) об адаптационно-трофической функции симпатической нервной системы. Ортопедическая неврология. Синдромология
Независимость трофического влияния нерва на мышцу от возбуждающих двигательных импульсов установлена в опытах инактивации мышц с помощью тенотомии, а также фиксации суставов (Chor Н., Dolkart R., 1936; Katpati G., 1968), изолирования отдела спинного мозга (Towwer, 1937), новокаиновых блокад нервного ствола (Gutmann E., 1962). Электронно-микроскопические исследования показали уменьшение диаметра мышечных миофибрилл при всех видах атрофии (Wechsler W., 1966). Как в денервированной, так и в тенотомированной мышце, кроме того, снижается активность фосфорилирования из-за замедления фермен-тативно-фосфорилазной и фосфогексокиназной деятельности (Телепиева В.И., 1955). Нейрогенный вариант атрофии отличается от атрофии при инактивации большим темпом потери веса и более быстрым уменьшением диаметра мышечных волокон (Muscatello V., Patriarca P., 1968); наблюдается более резкое падение концентрации карнозина и креатина и более значительное увеличение содержания глутаминовой кислоты, чем при тенотомии (Юдаев НА. и др., 1953). Свободные Н-группы, появляющиеся в дегене-рированной мышце, содержатся в меньшем количестве в тенотомированной (Попова М.Ф., 1956). Существенные различия обнаружены в содержании нуклеиновых кислот: значительное повышение уровня ДНК, отмечаемое в денервированной мышце, не происходит при тенотомии, что связывают с различиями в числе мышечных ядер (Hollos J. et al., 1960; Gutmann К, 1961, 1962). Содержание РНК в первые дни понижается больше в денервированной мышце, чем в тенотомированной (Hobos G. etal., 1960). Специфическим для денервированной мышцы является повышение ее чувствительности к медиаторам, особенно к ацетилхолину (Кеннон У., Розенблют Б., 1951), снижение уровня дыхания (Никитин В.Н. и др., 1956), повышение содержания воды, что не наблюдается в тенотомированных мышцах. Регулярная электростимуляция обездвиженных мышц предохраняет их от атрофии и поддерживает нормальную мышечную силу (Кеннон У., Розенблют А., 1951). Электротерапия денервированных мышц задерживает атрофию на определенный период времени, сила сокращения при этом уменьшается, повышается чувствительность к ацетилхолину (Гинецинский А.Г., 1956), содержание ДНК не меняется (Gutmann Z, 1961). Авторы оценивают это как доказательство наличия специфического трофического влияния нерва. Стало ясно, что нервная система регулирует метаболизм мышцы, воздействует на анаболические процессы. Трофическая функция нерва заключается в усилении послефунк-циональных восстановительных процессов. Об этом можно судить по тому, что в денервированной мышце после ее раздражения не усиливается процесс ресинтеза гликогена, не повышается синтез белков, не наблюдается усиления утилизации безбелкового азота и повышения потребления кислорода, как это происходит после раздражения нормальной мышцы. Согласно гипотезе Е.Gutmann (1961), трофическое влияние нерва осуществляется путем транспортировки с током аксоплазмы определенного вещества, вырабатываемого нейроном (см. главу 8). Р.П.Женевская (1974) изучала пластические процессы мышечной ткани в условиях регенерации нерва в период до начала сократительной реакции мышцы в ответ на раздражение нерва. Установив факт трофического влияния нерва на денервированную мышцу в тот период, когда в процессе регенерации еще не сформирован мионевральный синапс, автор приходит к следующему заключению: нерв в данных условиях, когда невозможна передача возбуждающего импульса (еще отсутствует мионевральный синапс), не функционирует как компонент нервной системы. Следовательно, обнаруженное трофическое влияние осуществляется самой тканью двигательного нерва. При оценке результатов собственных экспериментов автор вслед за другими подтвердила, что трофическое влияние нервной системы заключается в передаче иннервируемым тканям нервно-трофических метаболитов, действующих по типу веществ, осуществляющих эмбриональную индукцию. Обнаруженный исследователем факт задержки роста ау-тотрансплантатов и реиннервированных мышц в отсутствии проприоцепции дает основание считать, что подача нервно-трофических метаболитов мотонейронам контролируется системой обратных связей по афферентным путям, возможно, также с помощью передачи специфических веществ от мышечной ткани. Сейчас уже известно, что движение веществ по нерву осуществляется в обоих направлениях: обнаружен ток веществ не только от тела нейрона к периферии, но и от иннервируемой ткани к телу нейрона по микротрубочкам (Weiss J., 1969). Эти сведения связаны с довольно детально разработанным учением об аксоплаз-матическом токе. Таким образом, существуют данные о роли афферентных путей из мышцы, обратных связей, регулирующих подачу нервно-трофических метаболитов мотонейронам. В свете всего изложенного выше состояние кровоснабжения мышцы, ее обеспеченность кислородом приобретают новое значение. Отмеченная стадийность мышечно-тро-фических и нейродистрофических процессов заставляет нас вспомнить о стадийности сосудистых сдвигов в тех же тканях. Известно, что первыми при ишемии страдают нервно-мышечные аппараты (Ходос Х.Г., 1940) при одновременной высокой чувствительности и сосудистых рецепторов к кислородному голоданию (Долго-Сабуров Б.А., 1956; Куприянов В.В., 1955 и др.). Некоторые сведения о нейродистрофических сдвигах в условиях ишемизации тканей дали следующие эксперименты. С.С.Трач и А.Р.Радзиевский (1970) перерезали артериальные магистрали и вводили в мышцы АТФ. При этом возникавшие атрофии были значительно меньше, чем при перерезке сосудов без введения АТФ. То же касалось и изменений в нервных структурах. Если без введения АТФ ишеми-зация ведет к распаду нервных окончаний вплоть до их фрагментирования, при введении АТФ грубых реактивно-дистрофических нарушений не было. Отмечались лишь реактивно-регенеративные изменения: вздутия и утолщения нервно-мышечных пластинок и разрастания шванновских структур. В саркоплазме вблизи нервно-мышечной пластинки уменьшалось количество митохондрий. Т.к. АТФ эффективен при введении его в ничтожно малых дозах, он действует на мышцу не непосредственно, а через нервно-мышечные приборы. В этой связи следует помнить основополагающие данные В.А.Энгельгарта (1939): миозин разлагается АТФ, а последняя и ее ферменты делают миозиновые волокна более растяжимыми. Глава III. Методики вертеброневрологического обследования 11 Как ни велика роль гипоксии в описанных дистрофических процессах в мышечных и фиброзных тканях, влияния ее самой по себе недостаточно для объяснения нейромио-и нейроостеофиброза. В поисках других механизмов этих процессов было обращено внимание на то, что они развертываются: а) не только в связи с микротравматизацией (роль статико-динамических перегрузок при остеохондрозе); б) не только в связи с вазомоторными нарушениями; в) не только в связи с эфферентными импульсами, следующими по поврежденному корешку (роль компрессионных механизмов остеохондроза); г) не только в связи с эфферентными импульсами, следующими по здоровым нервным стволам из зон ирритации позвоночника и соответствующего квадранта тела (роль рефлекторных механизмов остеохондроза). Было установлено, что тонические и нейродистрофичес-кие нарушения в мышечно-фиброзных тканях возникают нередко под влиянием ряда экзогенных факторов, обусловливающих аутоиммунизацию. Антигенные свойства, «чуже-родность» могут приобрести не только ткани, формировавшиеся изолированно от лимфоидной системы, как, например, выпавшие пульпозные ядра межпозвонковых дисков, но также, при определенных условиях, денатурированные белки любых тканей, в том числе мышечных и фиброзных периферических нервов. Рассмотрение нейродистрофических изменений мышц невозможно без одновременного анализа соответствующих изменений их соединительнотканных элементов и соединительной ткани вообще. Миофиброз и нейроостеофиб-роз — это в такой степени связанные звенья одной патологической цепи, что можно считать целесообразным включение в понятие нейроостеофиброза и явления миофиброза. Не удивительно, что и другие терминологические попытки совершаются в направлении подобных же обобщений — например, миоэнтезиты... Перед тем как рассмотреть различные стороны патогенеза не только мышечно-тонических, но и миофиброзных дистрофических процессов и, в частности, их аутоиммунных механизмов, остановимся на некоторых особенностях соединительной ткани вообще. И.И.Мечников (1892) высказал важные взгляды о роли клеток соединительной ткани в формировании иммунитета. Эти идеи получили дальнейшее развитие в трудах А.А. Богомольца (1936) о системе соединительной ткани. Она несет опорно-трофическую функцию, состоит из клеток, межклеточного аморфного вещества и волокнистых структур. Новообразование основного вещества соединительной ткани и ее коллагеновых волокон связано с жизнедеятельностью фибробластов. Биохимики установили сложную структуру главнейшего белка ткани волокнистых структур — коллагена. Гистохимия позволила более углубленно изучить группу коллагеновых заболеваний. J.Nageott (1927, 1931), учитывая данные Эвальда о растворимости коллагена в слабокислой среде, а также способность его выпадать из раствора в форме фибрилл под влиянием солей, выдвинул биохимическую гипотезу образования коллагена. Согласно этой гипотезе, растворимый коллаген образуется из некоего белка крови, а затем осаждается под влиянием солей в коллаген. Гистологи разделяют данную концепцию, подчеркивая, что коллаген образуется из лреколлагена. Обе точки зрения примиряет А.А.Тустанов-ский (I960), установивший, что растворимый коллаген является биохимическим предшественником коллагена, и этот предшественник они совместно с В.И.Ореховичем (1952) назвали проколлагеном (не путать с преколлагеном биохимиков). Т.к. в фибриллогенезе участвуют коллагено-вые и неколлагеновые белки и мукополисахариды, коллаген рассматривают и как белково-мукополисахаридный комплекс (Jackson D. et ai, 1958). Морфологи, установив, что в коллагеногенезе первым этапом бывает преколлаген, своими методами не могли решить, пропитывается ли коллаген растворимым коллагеном или, деградируя, замещается вторым, или превращается в него биохимически. По точному замечанию АА.Тустановского, «...биохимики не изучали того, что видели морфологи, а морфологи не видели того, что предлагали видеть биохимики» (1960, с. 8). Г.В.Орловская и АЛ.Зайдес (1956) показали, что аргирофильный преколлаген эмбриональной закладки, не обладая рентгенострук-турными особенностями коллагена, со временем приобретает их. Оказалось, что белковая основа коллагена — комбинация двух белковых компонентов, один из которых — про-коллаген, другой — колластромин, который генетически связан с проколлагеном морфологов. Проколлаген и колластромин взаимодействуют по типу сочетания двух фаз: наружная фаза образована слоями проколлагена, внутренняя — нитями колластромина. В последних в дефинитивном коллагене сохраняется преколлаген как начальная фаза. Наименьшая структура единиц, содержащая обе фазы, — первичная коллагеновая фибрилла диаметром 500-1000 А. Эта единица одновременно морфологическая (на электронном уровне) и биохимическая. Каждая фаза содержит полисахариды, а колластроминовая — еще и глобулярные белки. Она занимает 80% свежего веса коллагена. Коллаген не идентичен волокну, он первичная коллагеновая фибрилла, элемент коллагенового волокна, но уже является многофазной и многокомпонентной системой (Зай-десА.Л., 1960). Образование компонентов коллагена, способы их сочетания остаются в области биохимии. Образование же коллагенового волокна — область фибриллогенеза. Коллагенообразование — стадия волокнообразования. Эластичные волокна также состоят из коллагеновых, только очень тонких, фибрилл. В них также имеются мукополисахариды. Кроме того, из них выделен белок эластин. Применительно к остеохондрозу важно, что образование коллагена нарушается при С-авитаминозе. McCormick (1954) подчеркивает, что при этом нарушается синтез коллагена в тканях диска, т.е. наступает дистрофия фиброзного кольца. Потребление оптимального количества витамина С является фактором сохранения целостности межпозвонкового диска (Greenwood J., 1964). В.Б.Киселев (1971) у больных остеохондрозом выявил С-витаминную недостаточность, характерную для них во все сезоны года. Недостаток аскорбиновой кислоты влияет на выработку проколлагена (Орехович В.Н., 1952) и полисахаридов, участвующих в его формировании. Важными для регуляции коллагенообразования и других функций соединительной ткани являются гормональные факторы, в первую очередь состояние системы гипофиз — кора надпочечников. Введение АКТГ, глюкокортикоидов, гормона щитовидной железы угнетает размножение фибробластов, задерживает развитие грануляционной ткани. Противоположное действие оказывает соматотропный гормон. Тестостерон стимулирует образование основного вещества и способствует аккумуляции в нем серы. 78 Ортопедическая неврология. Синдромология В старости известны атрофические изменения коллаге-новых волокон при увеличении эластина, в частности в коже. В основном веществе соединительной ткани количество кислых мукополисахаридов уменьшается, накапливаются мукопротеиды, нарушаются процессы диффузии и ухудшается питание клеток, прогрессируют дистрофические процессы. Такие изменения А.В.Мельченко и соавт. (1965) отмечали и в дистрофичных межпозвонковых дисках. В патологических условиях при бесклеточных склерозах (Абрикосов А.И., Струков А. И., 1953, 1954) вместо оформленных преколлагеновых волокон образуются аргирофиль-ные белково-полисахаридные массы, на которых неупорядоченно откладывается проколлаген. При разрастании соединительной ткани с участием клеток преколлагеновые ар-гирофильные волокна претерпевают преобразование по эмбриональному типу. При коллагеновых болезнях наблюдается дезорганизация коллагеновых волокон. Под разрешающим воздействием экзогенных факторов сами по себе денатурированные белки соединительной ткани могут стать внутренним источником реактивных, в частности, аутоиммунных процессов. Аллергический же фактор, как показали О.Я.Острый и З.И.Собиева (1961), А.И.Зайко (1970) и др., является одним из компонентов не-врогенной дистрофии. Другими словами, компонентом нейроостеофиброза являются и аутоиммунные реакции, включающиеся в нейрогенный дистрофический процесс при остеохондрозе. Этот процесс, таким образом, зависит и от особенностей взаимодействия аутоантигенов с соответствующими антителами. А.Д.Адо (1970) классифицирует антитела по трем категориям: 1) агрессивные (с цитотоксическим повреждающим действием на клетки и ткани); 2) аутоантитела-свидетели (без агрессивных свойств), наличие которых выявляется серологическими реакциями и указывает на факт воздействия аутоантигенов; 3) защитные аутоантитела, способствующие выздоровлению организма от аутоиммунного заболевания и переходу аутосенсибилизации в состояние резистентности к данному тканевому раздражителю. Они также транспортируют к местам выделения и обезвреживают постоянно возникающие продукты распада клеток совершенно здорового организма. 3.2.3.2. Субстраты, процессы и клинические проявления локальных мышечных гипертонусов Выше мышечные уплотнения были рассмотрены с точки зрения их возможной биохимической структуры. Рассмотрим их с точки зрения сократимости и контрактуры. Локальные мышечные гипертонусы — своеобразные локальные контрактуры. Понятие контрактуры применяется ортопедами для обозначения стойкого ограничения движения в суставе. Однако уже Figel (1876), который ввел это понятие в медицину, рассматривал его шире — как измененное состояние мышцы, ее стойкое укорочение. Причины этого укорочения могут гнездиться в самой мышце и аппаратах ее иннервации, включая рецепторные. Но и в последнем случае контрактуру усугубляют факторы непосредственного воздействия на мышцу. Контрактура легче возникает в красных мышцах, особенно в период утомления, при воздействии кислот, ток- сических веществ, при понижении температуры мышцы. В последнем случае меняются ее вазомоторные реакции на раздражение симпатикуса — дилатация вместо спазма (Родионов И.М., 1963). Контрактуры делят на активные, т.е. нейрогенные, и пассивные, т.е. фибротические, необратимые. Впрочем, длительно существующие активные контрактуры благодаря фиброзу также становятся пассивными. Но в них мы обнаруживаем некоторые электромиографические черты активных контрактур (Попелянский Я.Ю. и соавт., 1993). А.Г.Гинецинский (1945) в согласии с C.Sherrington (1907, 1935) считал, что в основе активных контрактур лежит рефлекс на растяжение, т.е. раздражение проприорецепторов мышцы и связанных с ними фиброзных образований. В последующем было установлено, что состояние проприорецепторов регулируется, настраивается импульсами, которые проводятся тонкими переднекорешковыми у-эффе-рентными волокнами (LexellL., 1945, 1956). При этом выявление контрактуры и ее выраженность зависят не только от афферентных и у-эфферентных импульсов, но и от состояния центральных аппаратов. Состояние это в русской физиологической литературе неоднократно определялось: парабиоз по Н.Е.Введенскому (1901), парабиотические очаги (Березина М.П., 1944), фиксация возбуждения в центрах (Орбели Л.А., 1945; Гинецинский А.Г., 1945), стационарное возбуждение в центрах (Уфлянд Ю.М., 1950), начальная стадия парабиоза с характерной для нее повышенной возбудимостью (Жуков Е.К., Думова A.M., Верещагин СМ., 1952). Эти проявления А.Г.Гинецинский (1945), Л.Н.Зефиров и В.А.Алатырев (1971) отождествляют с доминантой А.А.Ухтомского, т.к. центры во время доминанты находятся в стационарном парабиотическом возбуждении. Если оно продолжительно, говорят о патологической доминанте, и в этой связи это соответствует клиническим особенностям тех мышечных контрактур, которые мы наблюдаем при фиксированных кифозах, лордозах и сколиозах (исключая, возможно, альтернирующие). Н.Е.Введенский (1912) описал и иное состояние: при непрерывной в течение нескольких минут афферентной импульсации из одного чувствительного нерва повышается возбудимость другого нерва. Однако состояние это удерживается лишь несколько часов. При раздражении висцеральных нервов оно бывает более длительным (Айрапетьянц Э.Ш., 1957). Учитывая сказанное, можно сделать вывод, что различные контрактуры на фазах их развития реализуются преимущественно то по типу переходной фазы парабиоза, то по типу истериозиса. При оценке центральных механизмов контрактуры следует учитывать, во-первых, сегментарные влияния импульсных и аксоплазматических воздействий на мышцу. В нашей клинике было установлено, что и аксоплазматиче-ский ток по нерву в условиях повреждения мышцы может быть включен по рефлекторным путям, столь важным в патогенезе контрактуры (Попелянский Я.Ю., Хабиров Ф.А., 1991). Во-вторых же, надсегментарные аппараты усиливают активность и g-эфферентов, что, согласно J.Granit (1956), играет важную роль и в возникновении децеребрационной ригидности. Надсегментарные аппараты и, в особенности, вентромедиальные части ретикулярной формации оказывают и тормозящее влияние на сегментарную активность. Это обеспечивает торможение спинальных рефлекторных цент- Глава III. Методики вертеброневрологического обследования 79 ров в первой, латентной, фазе контрактуры, вызываемой патологической афферентацией. В условиях эксперимента эта фаза при перерезке спинного мозга укорачивается. При сохранении же супраспинальных влияний латентная фаза продолжительна (3-8 минут). Чем сильнее афферента-ция, тем продолжительнее это тормозное влияние. Лишь вслед за тем наступает вторая фаза повышенной возбудимости. Кроме перечисленных периферических и центральных нервных механизмов контрактуры, ее возникновению способствуют гуморальные факторы, в частности, наличие адреналина, активная функция мозгового слоя надпочечников (Кибяков А.В., Попов А.Ф., 1958; Волкова И.Н., Коч-нев О.С., 1967и др.), а также определенное состояние симпатической нервной системы. Оно сказывается на работоспособности скелетных мышц (феномен Гинецинского-Орбе-ли), на состоянии спинальных центров (Орбели Л.А., 1938; Лехтман Я.Б., 1969 и др.). Раздражение симпатических образований способствует появлению рефлекторной контрактуры, а удаление симпатических ганглиев — ее исчезновению (Еселевич Э.И., 1947; Четвериков П.С, 1968; Галиц-кая Н.А., 1965). Итак, в развитии активной контрактуры играют роль все элементы нервной системы от рецепторов и эффекторов ее и кончая сегментарными и надсегментар-ными нервами и нервно-гуморальными аппаратами. Клиническими и клинико-физиологическими исследованиями наших сотрудников показано, что важную роль в возникновении мышечно-тонического напряжения при остеохондрозе играет раздражение рецепторов чревных, тазовых, диафрагмальных и других вегетативных нервов при поражении внутренних органов (Попелянский Я.Ю., 1962; Петров Б.Г., 1965, 1969; Веселовский В.П., 1986; Богданов Э.И., 1989;Хабиров Ф.А., 1991 и др.). На основании общефизиологических положений можно считать, что при контрактуре осуществляется усиленная циркуляция импульсов, их дисперсия в межнейронных «путях запаздывания» и в нейронных кругах. Этим объясняют длительные следовые последействия в центрах. При оценке указанных циркуляции импульсов по нейронным кольцам (Самойлов А.Ф., 1930; Бернштейн Н.А., 1947 и др.) следует учитывать обратные сигналы и из самих мышц, а также особенности реализации контрактуры в зависимости от характера и состояния самих мышц. Мы попытаемся показать, что и клинические наблюдения подтверждают эти данные физиологов, но не при всех вариантах контрактур (см. ниже). Все описанные процессы в сегментарных и надсегмен-тарных нервных аппаратах меняют функциональное состояние самой мышцы, особенно в случаях ее преморбидной дефектности. В ней меняются процессы интимные («внутренние»), как сказано в приводимом ниже определении контрактуры. Интенсивность регистрируемых электрических процессов не всегда адекватна интенсивности укорочения. Согласно определению J.Travell, D.Simons (1983), контрактура — «устойчивая внутренняя активация: сокращение мышечных волокон происходит при отсутствии потенциалов действия в мотонейронах» (с. 2). Точность данного определения, как и практичность определения ортопедов, в свое время были одинаково оправданы. Развивающаяся вертеброне-зрология потребовала все же сделать два уточнения: 1) активная контрактура — отнюдь не одно лишь ограничение дзижений в суставе за счет фиброза, одним из важнейших ее Дата добавления: 2015-12-16 | Просмотры: 700 | Нарушение авторских прав |