|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Проверка модели Уотсона-КрикаМодель двойной спирали ДНК была предложена в 1953 г. и знаменовала рождение новой науки-молекулярной биологии. Однако прошло пять лет, прежде чем были получены первые убедительные экспериментальные подтверждения модели Уотсона-Крика в работах Мэтью Мезелсона и Франклина Сталя. В этих экспериментах было показано, что в точном соответствии с предсказаниями модели репликация ДНК происходит полу консервативно: каждая дочерняя молекула ДНК состоит из одной интактной (консервативной) цепи, полученной от родительской
4. Природа генетического материала 109
двойной спирали, и одной вновь синтезированной цепи. С другой стороны, можно представить себе гипотетические механизмы репликации ДНК, которые не предсказываются моделью двойной спирали, а именно: 1) консервативный способ репликации, при котором родительская ДНК полностью сохраняется, а дочерние молекулы ДНК полностью синтезируются заново, и 2) дисперсный способ, при котором обе дочерние молекулы ДНК синтезируются заново, а родительская молекула распадается на нуклеотиды, которые могут входить или не входить в состав дочерних молекул.
110 Организация и передача генетического материала
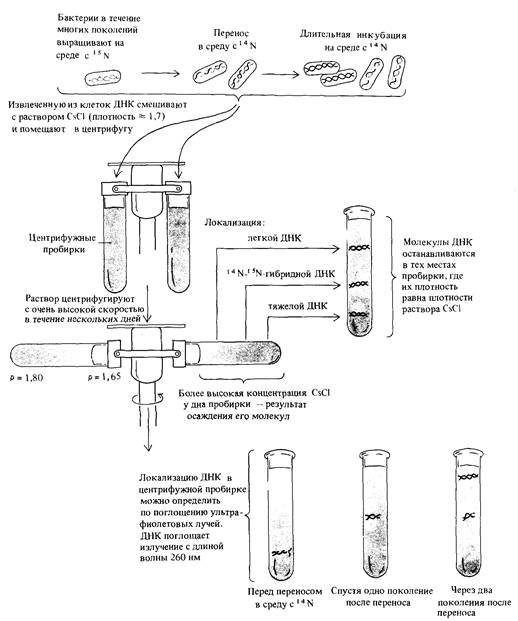
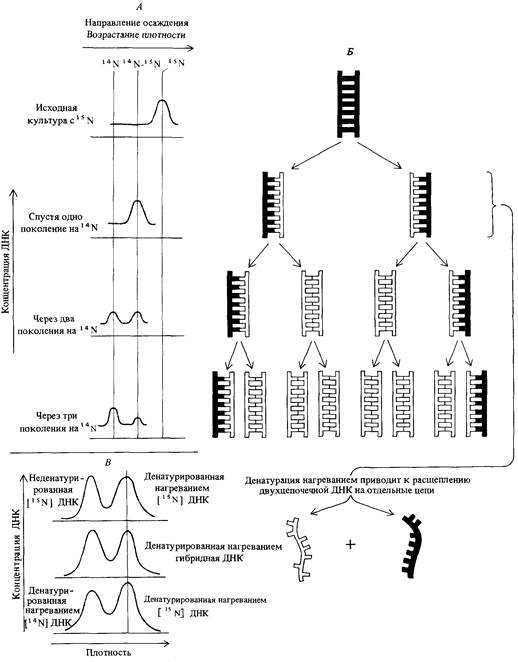
4. Природа генетического материала 111 Чтобы определить, каким из этих способов реплицируется ДНК. надо уметь отличать дочерние молекулы от родительских. Мезелсон и Сталь выращивали Е. coli на среде, содержащей в качестве источника азота 15N. Тяжелый изотоп азота 15N включался в состав ДНК и служил меткой. Для того чтобы пометить изотопом 15N практически всю бактериальную ДНК, достаточно вести культивирование на такой среде в течение двенадцати поколений. Молекулы, содержащие 15N, можно отличить от молекул, содержащих более легкий, обычный изотоп, по плотности, так как у ДНК с 15N масса одного нуклеотида больше, чем у обычной ДНК. Молекулы ДНК с различной плотностью могут быть разделены центрифугированием в градиенте плотности хлористого цезия (рис. 4.14). В процессе центрифугирования молекулы ДНК собираются в том слое, в котором плотность раствора равна их собственной плотности. ДНК клеток Е. coli, выращенных на среде, содержащей 15N, имеет плотность 1,724 г/см3, тогда как ДНК клеток, выращенных на обычной среде с изотопом 14N, имеет плотность 1,710 г/см3. Смесь этих двух типов ДНК легко разделяется при центрифугировании по плотности (рис. 4.14). Аналогией, иллюстрирующей принцип центрифугирования в градиенте плотности, может служить погружение подводной лодки. Регулируя количество воды в балластных танках (баках), подводная лодка может зависать на желаемой глубине, плотность воды на которой равна средней плотности подводной лодки. Если набрать в танки больше воды, вытеснив соответствующее количество воздуха, то плотность подводной лодки увеличивается, и она погружается, попадая в слои более холодной воды с большей плотностью. Когда плотность воды оказывается равной плотности подводной лодки, лодка снова зависает на новой глубине. В эксперименте Мезелсона и Сталя клетки, в течение многих поколений культивируемые на среде с 15N, быстро переносили в среду, содержавшую 14N. Через определенные промежутки времени отбирали пробы растущей культуры и в каждой из них определяли плотность ДНК. Результаты представлены на рис. 4.15. После первого деления на среде с 14N плотность ДНК культуры была промежуточной между [15N] ДНК и [14N] ДНК. После второго деления на среде с 14N половина клеток имела легкую ДНК, а вторая половина - ту же, что и в предыдущем поколении (промежуточную). После третьего деления на среде с 14N 3/4 ДНК имело плотность, равную плотности [14N] ДНК и 1/4 сохраняла промежуточную плотность. Соотношение между числом генераций и распределением плотности ДНК в точности соответствовало полуконсервативному типу репликации, предсказываемому моделью Уотсона-Крика (рис. 4.15). Кроме того, модель предсказывала, что ДНК с промежуточной плотностью должна представлять собой гибридную двойную спираль, одна из цепей которой содержит только тяжелый изотоп азота (N15), а другая-только легкий. Мезелсон и Сталь нагревали ДНК промежуточной плотности в течение 30 мин при температуре 100°С, что, как уже было известно, изменяет физические свойства молекулы, не разрывая ковалентных связей, и обнаружили, что она превращается в две равные по объему фракции ДНК с разными плотностями. Плотность одной из фракций, образовавшихся в результате нагревания, совпадала с плотностью тяжелой ДНК, а другой-с плотностью легкой ДНК (рис. 4.15).
112 Организация и передача генетического материала
4. Природа генетического материала 113 Из этого было сделано заключение, что ДНК промежуточной плотности, образующаяся после первого деления в легкой среде, представляет собой гибридную молекулу, состоящую из одной родительской цепи, содержащей только тяжелый изотоп азота, и другой, вновь синтезированной дочерней цепи, как это предсказывает модель Уотсона-Крика. Аналогичные эксперименты проделывали с реплицирующейся ДНК множества различных прокариотических и эукариотических организмов, и каждый раз оказывалось, что ДНК реплицируется полуконсервативно. Эксперименты Мезелсона-Сталя были первым доказательством справедливости модели Уотсона - Крика. В настоящее время можно с уверенностью сказать, что основные положения этой модели убедительно подтверждены и структура двойной спирали легла в основу современной генетики. Дата добавления: 2015-12-16 | Просмотры: 708 | Нарушение авторских прав |
 Рис. 4.13. Предложенная Уотсоном и Криком модель репликациидвойной спирали ДНК.
Рис. 4.13. Предложенная Уотсоном и Криком модель репликациидвойной спирали ДНК.