|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Рекомбинация сцепленных геновОбъединение множества генов в одной хромосоме определяет характер наследования признаков, контролируемых этими генами. Гены, находящиеся в одной хромосоме, часто не расходятся независимо и потому представляют собой второе исключение из законов Менделя (первым было рассмотренное нами ранее наследование, сцепленное с полом). Гены, характер наследования которых отличается от независимого расщепления, называются сцепленными. Во второй главе мы видели, что Мендель постулировал два возможных исхода скрещивания двух чистых линий гороха, одна из которых имеет гладкие желтые семена (RR Y Y), a другая - морщинистые и зеленые (rrуу). Если при формировании гамет в поколении F1 распределение аллелей независимо, то следует ожидать возникновения в равных долях четырех типов гамет (RY,Ry,rY и ry), что ведет к появлению в поколении четырех фенотипов в отношении 9:3:3:1. Если же аллели каждого родителя оставались при формировании гамет в поколении f1 вместе, то возникли бы лишь два типа гамет: RY и rу. Другими словами, гены, определяющие форму семян и их цвет, были бы полностью сцеплены. Результат такого полного сцепления проявлялся бы в поколении F2. Оно было бы представлено лишь растениями двух типов с гладкими желтыми семенами и с морщинистыми зелеными в отношении 3:1. Если бы эти гены были полностью сцеплены, то каждая
130 Организация и передача генетического материала
пара признаков (гладкие и желтые, а также морщинистые и зеленые) наследовалась бы как один признак. Генетики предпочитают изучать сцепление посредством анализирующего скрещивания, т. е. скрещивания с родительской линией, гомозиготной по рецессивным генам (рис. 5.2). При анализирующем скрещивании фенотип потомства прямо отражает типы гамет, формируемые гетерозиготным родителем, как это показано для случаев независимого расщепления и полного сцепления на рис. 5.3. В первом из представленных на рис. 5.3 случаев при независимом расщеплении аллелей двух генов следует ожидать, что в равном отношении образуются четыре генотипа. Два из них содержат те же сочетания аллелей, что и у родителей (а + b + и ab), a два - новые рекомбинантные сочетания аллелей (а + b и ab +). Если в потомстве родительские типы и рекомбинантные типы представлены в равном отношении, то, значит, гены α и b в мейозе гетерозиготного родителя расходятся независимо и могут быть названы несцепленными. Частота рекомбинации между двумя генами определяется как доля обоих рекомбинантных типов в потомстве (в примере, представленном на рис. 5.3, 50/100 = 0,5, или 50%). Если гены находятся в независимо расходящихся разных хромосомах, то частота рекомбинации равна 50%. Обратное, как мы вскоре увидим, вообще говоря, не всегда справедливо. Во втором из представленных на рис. 5.3 случаев абсолютная связь между двумя генами ведет к тому, что в потомстве от анализирующего скрещивания следует ожидать появления лишь двух генотипов. Такое полное сцепление может служить веским, хотя и не исчерпывающим свидетельством в пользу того, что оба гена расположены в одной хромосоме. На рис. 5.3 приведен пример анализирующего скрещивания, выявляющего полное сцепление двух рецессивных аутосомных признаков
5. Геном эукариот 131
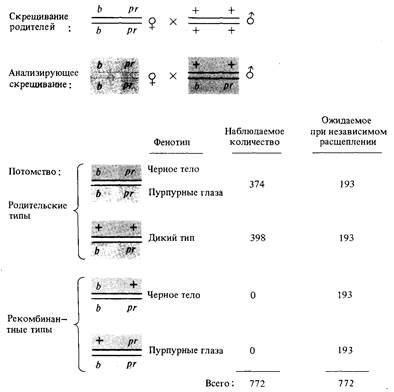
дрозофилы: черный цвет тела (b) и пурпурные глаза (рr). Если самец гетерозиготен по обоим генам, то комбинации аллелей в спермиях тождественны их комбинации в родительском генотипе, в результате чего в потомстве у половины мух тело черное и глаза пурпурные, а вторая половина потомства принадлежит дикому типу. Рекомбинантных сочетаний родительских аллелей не наблюдается: ни черных мух с нормальными глазами, ни мух с нормальным цветом тела и пурпурными глазами в потомстве нет. Представленные на рис. 5.4 данные указывают на полное сцепление генов, изучавшихся в этом скрещивании, из чего следует, что оба гена расположены в одной и той же хромосоме. Полного сцепления между аллелями двух различных генов обычно не наблюдается. Действительно, если признаки «черное тело и пурпурные глаза» всегда полностью сцеплены, т. е. всегда в мейозе расщепляются вместе, то мы вправе предположить, что оба признака являются проявлениями одного мутантного гена. Причину, по которой эти два гена ведут себя при скрещивании, представленном на рис. 5.4, как полностью сцепленные, мы обсудим после того, как рассмотрим результаты аналогичного скрещивания, в котором самка гетерозиготна по обоим генам. В этом случае, как показано на рис. 5.5, в потомстве представлены четыре типа: два родительских (черное тело, пурпурные глаза и дикий тип) и два рекомбинантных (черное тело, нормальные глаза и нормальное тело, пурпурные глаза). Это те четыре типа, которых следует ожи-
132 Организация и передача генетического материала
дать, если анализируемые гены расходятся независимо. Однако мух рекомбинантных типов много меньше, чем мух, имеющих родительский генотип (30/450), тогда как при независимом расхождении их число должно было бы быть примерно одинаковым (240/240). Такое отклонение от ожидаемого при независимом расщеплении свидетельствует о наличии сцепления. Генетики оценивают степень сцепления в таком скрещивании частотой рекомбинантных типов в потомстве: частота рекомбинаций в этом случае равна 30/480 = 0,0625, или 6,25%. Частота рекомбинаций для несцепленных генов равна 50% (240/480 = 0,50). Рекомбинантные типы при таком скрещивании (рис. 5.5) возникают из рекомбинантных гамет, образующихся у самки в процессе кроссинговера при мейозе. Кроссинговер обычно происходит в мейозе всех организмов, и у самцов, и у самок, и во всех парах гомологичных хромосом. Однако у самцов многих видов насекомых отряда Diptera, в том числе и у дрозофил, Кроссинговер не происходит и рекомбинантные гаметы не возникают. Вот почему в скрещивании, представленном на рис. 5.4, аллели генов, находящихся в одной хромосоме, расходятся всегда вместе, т.е. ведут себя так, как этого следует ожидать при полном сцеплении. Рис. 5.6 знакомит нас с примером наследования двух сцепленных генов Drosophila melanogaster, находящихся в Х-хромосоме. Все мужское потомство получает свою единственную Х-хромосому от матери, и эта Х-хромосома может быть как родительского, так и рекомбинантного типа. В этом примере анализирующее скрещивание гетерозиготной по обоим генам самки с самцом-носителем рецессивных аллелей обоих ге-
5. Геном эукариот 133
нов не обязательно, поскольку частоту рекомбинации можно определить по фенотипам гемизиготного мужского потомства. В Y-хромосоме нет аллелей соответствующих генов, расположенных в Х-хромосоме. В этом скрещивании двумя маркерами служили ген желтой окраски тела (у) и ген белоглазия (w); соответствующие доминантные аллели дикого типа определяют коричневатый цвет тела (у +) и красные глаза (w +). Частота рекомбинации между этими генами, наблюдавшаяся при таком скрещивании, составляла 4/3 90 x0,0 10. Анализ частоты рекомбинаций может дать ответы на два вопроса. Первый вопрос состоит в том, принадлежат ли оба гена одной хромосоме. Если наблюдаемая частота рекомбинаций между аллелями какихлибо двух генов меньше 50%, то на этот вопрос можно ответить положительно. Для генов, расположенных в одной хромосоме очень близко друг к другу, наблюдается обычно очень низкая частота рекомбинаций. По мере увеличения расстояния между генами частота рекомбинаций также увеличивается. Более того, кроссинговер между генами, находящимися в одной хромосоме, но сильно удаленными друг от друга, может происходить настолько часто, что наблюдаемая частота рекомбинаций будет близка к 50%, т.е. к значению, соответствующему независимому расщеплению. В таких случаях для определения принадлежности генов одной хромосоме требуется статистический анализ. Иногда хромосомы бывают настолько длинными, что гены, расположенные на разных концах, всегда расходятся независимо. Такие случаи можно выявить, лишь ответив на второй вопрос: каково взаимное расположение сцепленных генов в хромосоме? Рекомбинационный анализ дает ответ и на этот вопрос.
134 Организация и передача генетического материала Дата добавления: 2015-12-16 | Просмотры: 1465 | Нарушение авторских прав |