|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Внеядерная наследственностьПринципы генетического анализа применялись при исследовании многих эукариотических организмов: высших животных и растений, а также более просто устроенных эукариот, таких как грибы, водоросли и простейшие. Составлены подробные хромосомные карты множества различных видов, представляющих научный или практический интерес. Эти карты детально описывают организацию генов в ядрах клеток. Однако не все эукариотические гены локализованы в хромосомах клеточных ядер. Вскоре после переоткрытия менделевских законов наследования стало ясно, что некоторые типы изменчивости не подчиняются этим законам. В 1909 г. Карл Корренс опубликовал работу по наследованию пестролистности у Mirabilis jalapa, в которой был описан неменделевский тип наследования. Для многих видов декоративных растений характерна пестролистность - появление белых или желтых пятен и полос на листьях зеленых растений (рис. 5.21). Желтые участки могут быть небольших размеров,
5. Геном эукариот 151
однако иногда желтыми становятся целые побеги, тогда как другие остаются зелеными или пестрыми. Корренс брал пыльцу с цветков, растущих на желтых, пестрых и зеленых побегах, и наносил ее (предварительно удалив тычинки) на пестики цветов, растущих на побегах всех трех типов. Оказалось, что свойства проросших из таких семян растений определяются исключительно характером материнского цветка и не зависят от свойств цветка, с которого была взята пыльца (табл. 5.2). Эти результаты были первым примером внеядерной, или цитоплазматической, наследственности. В настоящее время мы знаем, что зеленый цвет растений определяется хлоропластами, содержащими фотосинтетический пигмент хлорофилл. Зеленые хлоропласты развиваются из самостоятельно делящихся органелл, называемых пластидами, и находящихся в цитоплазме клеток растений. Пластиды клеток из желтых участков пестролистных растений не способны развиться в нормальные зеленые хлоропласты. Растения, выросшие из семян, завязавшихся в цветках на желтых побегах (табл. 5.2), не способны к фотосинтезу и скоро погибают. Пестролистность растения является следствием присутствия в цитоплазме зародыша обоих типов самореплицирующихся пластид. В каждой клетке растения содержится сравнительно небольшое число пластид. По мере роста растения посредством клеточных делений некоторые дочерние клетки получают случайно лишь нормальные пластиды, другие - лишь неспособные к фотосинтезу, третьи - смесь и тех и других. Эти клетки дают начало соответственно зеленым, желтым и пестролистным побегам. Представленные в табл. 5.2 результаты показывают, что пластиды семян наследуются от материнской цитоплазмы, а не от пыльцы. Этот вывод впоследствии был подтвержден микроскопическими наблюдениями. Способностью к саморепликации лишенные хлорофилла пластиды (а значит, и нормальные пластиды) обязаны содержащейся в них ДНК, в которой генетически закодированы функции, необходимые для нормальной жизнедеятельности пластид. ДНК выделена из хлоропластов растений многих видов. Ее выделение часто облегчается тем, что ну-
152 Организация и передача генетического материала
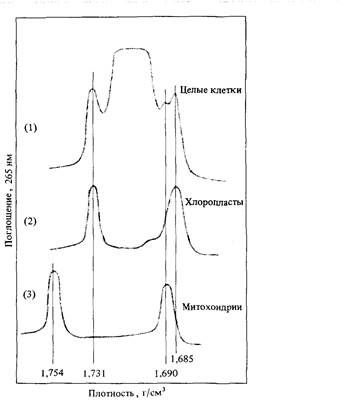
клеотидный состав ДНК многих хлоропластов сильно отличается от состава ядерной ДНК, и, следовательно, эти две фракции легко разделяются при центрифугировании в градиенте хлористого цезия. Например, ДНК хлоропластов фотосинтезирующих одноклеточных водорослей Euglena glacilis имеет плотность 1,685 г/см3, и ее можно легко идентифицировать как сателлит ядерной ДНК, обладающий более высокой плотностью (рис. 5.22). В утративших хлоропласты линиях того же вида ДНК с соответствующей плотностью отсутствует. Выделенные из хлоропластов Euglena glacilis молекулы ДНК имеют кольцевую форму и длину 132000 нуклеотидных пар. Хлоропласты обладают своей собственной системой синтеза белка, сильно отличающейся от соответствующей системы клеточной цитоплазмы. В них также содержатся пигменты, ферменты и другие белки, необходимые для синтеза углеводов из углекислого газа и воды при участии солнечного света (фотосинтеза). Некоторые из генетических функций, необходимых для фотосинтеза и синтеза белков хлоропластами, закодированы в ДНК хлоропластов, другие - в ядерной ДНК. Механизмы фотосинтеза и другие функции хлоропластов активно исследуются посредством генетического анализа мутаций, затрагивающих эти функции. Некоторые из этих мутаций при скрещиваниях обнаруживают менделевское расщепление и, следовательно, относятся к генам ядерной ДНК. Для других характерно неменделевское наследование; следовательно, соответствующие гены локализованы в хлоропластах.
5. Геном эукариот 153 В выделенной из Euglena ДНК присутствует еще одна сателлитная фракция, сохраняющаяся в клетках, лишенных хлоропластов (рис. 5.22). Этот тип ДНК локализован в митохондриях - органеллах, ответственных за дыхательную активность клеток. В митохондриях в процессах окислительного фосфорилирования и в цикле трикарбоновых кислот образуется большая часть АТР (аденозинтрифосфата) клетки. АТР-это основной источник энергии, расходуемой на все метаболические реакции клетки. Митохондрии, так же как и хлоропласты, способны к саморепликации. В их ДНК закодировано множество функций, необходимых для нормальной дыхательной активности. Литература Дата добавления: 2015-12-16 | Просмотры: 1007 | Нарушение авторских прав |