|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Ключевые слова и понятия. Вирулентный фаг Гетероаллели Делеционное картирование Комплементационный тест Мутон Непермиссивные (рестриктивные) условия
| Вирулентный фаг
Гетероаллели
Делеционное картирование
Комплементационный тест
Мутон
Непермиссивные (рестриктивные) условия
Пермиссивные условия
Псевдоаллель
Расстояние на карте
Рекомбинационный тест
| Рекон
Сбалансированная деталь
Тонкая структура гена
Точечная мутация
Транс -конфигурация
Умеренный фаг
Условно летальная мутация
Цис -конфигурация
Цистрон
rII -система
|
Задачи
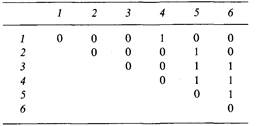
6.1. Обнаружено шесть новых мутаций фага Т4. С помощью комплементационного теста проведен попарный анализ всех мутаций. Результаты представлены в таблице. Знак «+» означает наличие прозрачного пятна (лизировавшие бактерии). Определите группы комплементации.
 6.2. У дрозофилы существует много рецессивных мутаций, обусловливающих стерильность самок, в частности это мутации dunce и diminutive. Они сцеплены с полом и на карте расположены очень близко друг от друга. Продумайте постановку эксперимента, позволяющего определить, аллельны ли эти мутации.
6.2. У дрозофилы существует много рецессивных мутаций, обусловливающих стерильность самок, в частности это мутации dunce и diminutive. Они сцеплены с полом и на карте расположены очень близко друг от друга. Продумайте постановку эксперимента, позволяющего определить, аллельны ли эти мутации.
| 6.3. Новая rII -мутация фага Т4 дает рекомбинанты дикого типа при скрещивании с у3 (см. рис. 6.10), но не дает их при скрещивании с 1241. Рекомбинанты дикого типа возникают также при скрещиваниях с 1993, 1695, РТ153 и Н88, но не появляются при скрещивании с 386 и 168. В каком участке карты rII локализована мутация?
6.4. rII -мутация, отобранная при длительном культивировании на среде, содержащей 5-бромурацил, дает рекомбинантов дикого типа при скрещивании с 638 (см. рис. 6.10). Кроме того, рекомбинанты дикого типа обнаруживаются в скрещиваниях с 1589 и 1299 и не обнаруживаются в скрещиваниях с 196, W8-33, 187 и 1519. Объясните результаты.
6.5. Картирование тонкой структуры с помощью делеций позволяет однозначно линейно упорядочить мутации. Отмечать при этом требуется лишь появление (или отсутствие) рекомбинантов дикого типа. В таблице представлены результаты попарных скрещиваний между
|
186 Организация и передача генетического материала
шестью rII -делениями; 1 означает появление рекомбинанта дикого типа при скрещивании, 0 - отсутствие. Постройте генетическую карту с указанием относительной длины каждой делеции. Какие из этих шести мутаций могли бы быть точечными и при этом результаты, представленные в таблице, не изменились?
 6.6. Альбинизм у человека обусловлен гомозиготностью по аутосомному рецессивному гену. В Англии описан случай, когда у супругов-альбиносов родились трое детей с нормальной пигментацией. Этот факт можно объяснить по крайней мере двояко. Как?
6.7. rII -Делеции, с помощью которых Бензер разбил область rII на 47 участков, были выбраны им из много большего набора rII -мутаций (рис. 6.10). Как и в условии задачи 6.5, расположение этих делеции друг относительно друга можно было установить, проводя попарные скрещивания и отмечая лишь присутствие или отсутствие рекомбинантов ди-
6.6. Альбинизм у человека обусловлен гомозиготностью по аутосомному рецессивному гену. В Англии описан случай, когда у супругов-альбиносов родились трое детей с нормальной пигментацией. Этот факт можно объяснить по крайней мере двояко. Как?
6.7. rII -Делеции, с помощью которых Бензер разбил область rII на 47 участков, были выбраны им из много большего набора rII -мутаций (рис. 6.10). Как и в условии задачи 6.5, расположение этих делеции друг относительно друга можно было установить, проводя попарные скрещивания и отмечая лишь присутствие или отсутствие рекомбинантов ди-
|  кого типа в потомстве. В таблице представлены результаты таких скрещиваний для 15 использованных Бензером неревертирующих мутаций. Постройте генетическую карту для этих мутаций. Считайте, что каждая мутация означает отсутствие одного определенного участка - другими словами, двойных мутаций в этой группе нет. (Указание: сначала определите порядок крупных делеций.)
6.8. Сбалансированные летальные линии сыграли большую роль в развитии генетики дрозофилы. Они сделали возможным конструирование сбалансированных хромосом, содержащих множественные инверсии, запирающие кроссинговер, рецессивные летали и гены стерильности самок, а также доминантные мутации, свидетельствующие о присутствии хромосомы в гетерозиготном состоянии. Так, например, Х-хромо-
кого типа в потомстве. В таблице представлены результаты таких скрещиваний для 15 использованных Бензером неревертирующих мутаций. Постройте генетическую карту для этих мутаций. Считайте, что каждая мутация означает отсутствие одного определенного участка - другими словами, двойных мутаций в этой группе нет. (Указание: сначала определите порядок крупных делеций.)
6.8. Сбалансированные летальные линии сыграли большую роль в развитии генетики дрозофилы. Они сделали возможным конструирование сбалансированных хромосом, содержащих множественные инверсии, запирающие кроссинговер, рецессивные летали и гены стерильности самок, а также доминантные мутации, свидетельствующие о присутствии хромосомы в гетерозиготном состоянии. Так, например, Х-хромо-
|
6. Тонкая структура гена 187
| сома, обозначаемая как FМ7, - это сбалансированная хромосома, содержащая инверсии, мутацию женской стерильности и доминантную мутацию Ваr(В), влияющую на форму глаз. Объясните, почему использование сбалансированной хромосомы позволяет неограниченно долго сохранять в Х-хромосоме летальные и другие вредные мутации?
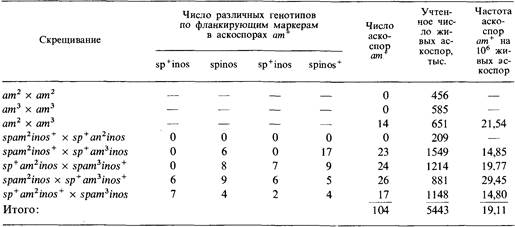
6.9. Мутации локуса pan y нейроспоры делают невозможным рост на среде, в которой отсутствует пантотеновая кислота, а мутанты по локусу trp могут культивироваться лишь в присутствии триптофана. Мутация ylo определяет желтый цвет спор (при нормальном черном). На основе представленных в таблице данных постройте карту, показывающую отношения между генами ylo и trp и различными мутациями локуса pan. Аскоспоры, полученные в этих четырех скрещиваниях, высевали на минимальную среду с добавлением лишь триптофана. Во всех скрещиваниях учитывали генотипы по неселективным маркерам среди рекомбинантов pan +.
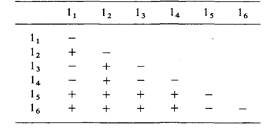
| 6.10. Отобраны шесть рецессивных летальных мутантов во второй хромосоме дрозофилы. Каждая линия сбалансирована SM1 (см. задачу 6.8) и маркирована доминантной мутацией Curly. Для того чтобы определить, являются ли какие-либо из этих деталей мутациями одного гена, были поставлены скрещивания типа ia/су ♀ x lB/Су ♂.
В приведенной ниже таблице знак «+» означает появление в потомстве особей, не несущих признак Су, знак «—» отсутствие таких особей. Мутациями скольких разных генов являются отобранные шесть деталей?
|
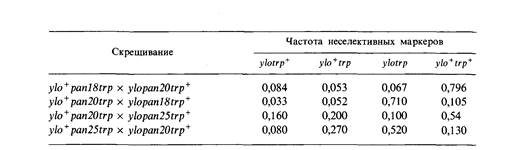
| | 6.11. У дрозофилы существует группа рецессивных летальных мутаций Minute (M), вызывающих замедленное развитие и утоньшение щетинок. При изучении мутаций Minute второй аутосомы самцов, гомозиготных по мутации cinnabar (cn), определяющей ярко-красный цвет глаз, подвергали рентгеновскому облучению и скрещивали с самками, у которых одна вторая хромосома несет сбалансированную деталь и маркирована доминантной мутацией Curly (Су) (загнутые крылья), а вторая- доминантной мутацией Brown (Bw). Потомков с загнутыми вверх крыльями, у которых, кроме того, были тонкие щетинки, скрещивали, как показано ниже, для того, чтобы выделить линию мух, несущих индуциро-
|
188 Организация и передача генетического материала
ванные Minute:
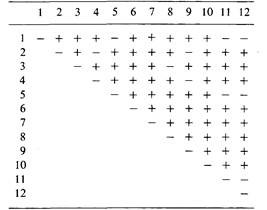
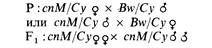 Каждое такое скрещивание давало линию, содержащую предполагаемую мутацию Minute. Истинные мутации Minute -рецессивные летали и идентифицируются по отсутствию гомозигот сп в потомстве от скрещивания F1. В описываемом эксперименте было обнаружено 12 вновь индуцированных мутаций Minute, каждая в сбалансированной летальной линии. Для того чтобы определить, относятся ли эти мутации к одному или различным генам, был поставлен тест на комплементацию. Для этого попарно скрещивали различные линии (пронумерованные от 1 до 12) и наблюдали за появлением в потомстве жизнеспособных гомозигот сп. Результаты скрещиваний представлены в таблице; «+» означает присутствие в потомстве жизнеспособных гомозигот сп, «—» означает их отсутствие. Разбейте эти 12 мутаций на группы комплементации.
Каждое такое скрещивание давало линию, содержащую предполагаемую мутацию Minute. Истинные мутации Minute -рецессивные летали и идентифицируются по отсутствию гомозигот сп в потомстве от скрещивания F1. В описываемом эксперименте было обнаружено 12 вновь индуцированных мутаций Minute, каждая в сбалансированной летальной линии. Для того чтобы определить, относятся ли эти мутации к одному или различным генам, был поставлен тест на комплементацию. Для этого попарно скрещивали различные линии (пронумерованные от 1 до 12) и наблюдали за появлением в потомстве жизнеспособных гомозигот сп. Результаты скрещиваний представлены в таблице; «+» означает присутствие в потомстве жизнеспособных гомозигот сп, «—» означает их отсутствие. Разбейте эти 12 мутаций на группы комплементации.
| 6.12. С помощью метода, схематически изображенного на рис. 6.2, определяли количество рекомбинантов между rII мутантами 165 и 201. При подсчете оказалось, что число фаговых частиц в пермиссивных условиях составляет 2000, а в непермиссивных - 50. Каково расстояние между 165 и 201 на генетической карте?
6.13. Самок, гетерозиготных по двум аллелям гена white (wa/wbf), скрещивали с самцами, гемизиготными по гену white. Из 100000 самцов в потомстве от таких скрещиваний лишь два были с глазами дикого типа. Каково расстояние между wa и wbf на генетической карте?
6.14. Сцепленная с полом доминантная мутация Notch (N) выражается в зазубренности крыльев, утоньшении жилок крыла и мелких неправильностях щетинок. Кроме того, эта мутация рецессивная леталь. Рецессивные мутации facet (fa) и split (spl) вызывают нарушения на поверхности глаз, но легко отличимы друг от друга. У мух с генотипами fa/N и spl/N проявляются признаки, характерные для соответствующих рецессивных мутаций и для генотипа N/ +; это означает, что N -мутантная хромосома не содержит аллелей fa + или spl +. Рецессивные мутации facet-notchoid (fano) и notchoid (nd) вызывают такие же нарушения в развитии крыльев, как и N. У мух с генотипом nd/N крылья зазубрены очень сильно, а генотип fa"°/N почти полностью летален; N -мутантная хромосома не содержит аллелей nd + и fano +. На основе приведенных ниже данных постройте карту этого псевдоаллельного локуса. В таблицу включены данные лишь по двум из многих N аллелей.
|
6. Тонкая структура гена 189
| Скрещивание
| Рекомбинантные самцы
| Общее число самцов
| | 
| |
|
|
|
|
|
|
|
|
|
|
| | По Welshons W. (1958). Ргос. Natl. Acad. Sei USA, 44, 254.
| | | По Welshons W., Halle Van E. (1962). Genetics, 47, 743.
| |
| 6.15. Как измерить частоту рекомбинаций между r168 и r924 при скрещивании r168r1695 x r1695r924 в ситуации, схематически изображенной на рис. 6.5? (Необходимо использовать простой и быстрый способ скрещивания, изображенный на рис. 6.11. Считайте, что вы располагаете линиями с единичными rII мутациями, а также двойными мутантами.)
6.16. Мутации нейроспоры am2 и αm3 делают ее неспособной к росту на среде
| без глицина или содержащей его в концентрации, меньшей 0,02 М. Этим можно воспользоваться в качестве инструмента для отбора рекомбинантов дикого типа, образующихся при скрещивании двух мутантных линий. Результаты скрещиваний представлены ниже; inos и sp- фланкирующие маркеры. Что можно сказать по результатам скрещивания?
|
Геном вируса
Вирусы - это сложные нуклеопротеины, использующие метаболический аппарат зараженной ими клетки для собственного размножения. Многие бактериофаги (например, Т4), вирусы растений и животных убивают инфицированные клетки в процессе размножения. Другие вирусы не разрушают полностью инфицированную клетку, а позволяют ей расти и делиться, производя и выделяя наружу потомство вирусов. Третьи, подобные бактериофагу λ и обезьяньему вирусу 40 (SV40), могут встраиваться в геном хозяина и пассивно реплицироваться по мере роста и деления клетки (т.е. переходить в состояние провируса).
Исследование вирусов, особенно бактериальных, внесло огромный вклад в наше понимание генетических явлений. Быстрое размножение бактериофагов дает возможность за одни сутки производить скрещивания в потомстве двух последовательных поколений. Аналогичные скрещивания на дрозофиле требуют 3,5 недель, а на кукурузе - по меньшей мере года. Кроме того, огромная численность фаговых популяций, содержащихся в нескольких миллилитрах культуральной жидкости, дает возможность наблюдать очень редкие генетические события. Малый размер геномов многих фагов по сравнению с геномом бактерий, например Е. coli, позволяет идентифицировать все или по крайней мере большинство фаговых генов и весьма подробно представить себе генетическую организацию и регуляцию генома в целом. Геном фага фХ174 состоит всего из девяти генов, геном фага лямбда - менее чем из 60, тогда как геном Е. coli насчитывает, вероятно, несколько тысяч генов. Сочетание этих замечательных достоинств сделало вирусы незаменимыми генетическими объектами и привело к тому, что геномы некоторых бактериофагов изучены в настоящее время лучше, чем каких бы то ни было иных организмов. Они могут служить моделями при анализе строения и работы более сложных геномов.
7. Геном вируса 191
Дата добавления: 2015-12-16 | Просмотры: 923 | Нарушение авторских прав
|