|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Бактериальный геномГенетические исследования организации генома бактерий начались вскоре после того, как было показано, что именно ДНК является веществом наследственности у пневмококков. Бактерии, так же как и вирусы, представляют генетикам возможность работать с популяциями колоссальной численности, затрачивая на эксперимент сравнительно небольшое время. Описываемые в этой главе методы отбора позволяют выявлять и изучать очень редкие генетические события. Объектом наиболее обширных и тщательных исследований служили и продолжают служить кишечные бактерии Escherichia coli и именно на них мы сосредоточим внимание в этой главе. Генетические свойства Е. coli характерны не только для этого вида бактерий, а методология генетических исследований, разработанная на Е. coli, создает фундамент и для изучения других видов. Исследования генетики бактерий внесли очень большой вклад в наши знания о наследственности. Во-первых, они продемонстрировали сколь разнообразны генетические процессы, которые могут реализовываться в природе у отдельных видов организмов. Познание этого разнообразия у прокариот проливает свет на возможные механизмы взаимодействия генома человека с геномами вирусов и приводит к переоценке роли многих генетических явлений, наблюдавшихся у эукариотических организмов, но не находивших объяснения. Велика роль генетики бактерий и в изучении регуляции и экспрессии активности генов. Эта тема будет рассматриваться в последующих главах. Механизмы организации этих процессов у сравнительно простых прокариотических организмов закладывают основы для их понимания у более сложно устроенных эукариот.
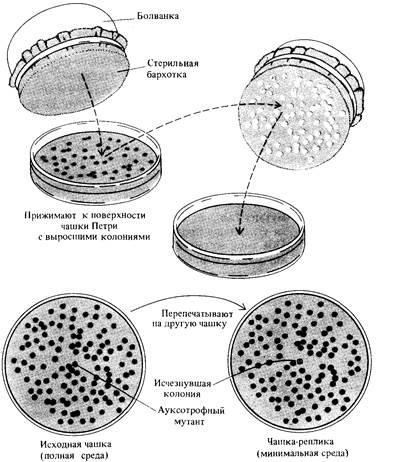
228 Организация и передача генетического материала Как отмечалось в гл. 7, генетический анализ генома вирусов формально развивался аналогично генетическому анализу, используемому при исследовании организмов, имеющих мейоз. Однако, когда этот же подход был использован для анализа мутаций Е. coli, возникло множество затруднений, пока генетики не осознали, что никакая аналогия между половым процессом у мейотических организмов и у бактерий невозможна. В настоящее время существуют представления, согласно которым бактерия содержит множество генетических элементов, более или менее независимых друг от друга и взаимодействующих между собой посредством механизмов, не находящих формальной аналогии с процессом мейоза. Открытие класса генетических элементов, названных эписомами, (в особенности F-эписом) и трансдуцирующих фагов дало возможность успешно применить принципы генетического анализа к бактериям и весьма подробно описать организацию бактериального генома. Мутанты Е. coli Прежде чем обсуждать генетику бактерий, мы должны познакомиться с типами изучаемых мутаций и с используемыми для них обозначениями. Е. coli дикого типа растет в лабораторных условиях на очень простой среде, единственным органическим составляющим которой служит источник углерода; как правило это глюкоза. Штаммы дикого типа прототрофны (см. главу 4): они способны синтезировать любые сложные органические молекулы, необходимые для их метаболизма и роста. Эти биосинтетические способности (анаболические функции) требуют работы (экспрессии) многих существенных (т.е. необходимых для существования бактерий) генов. Многие мутации, нарушающие экспрессию необходимых биосинтетических функций, называются условно летальными (см. главу 7), поскольку бактерии с такими мутациями могут существовать только при добавлении в среду необходимых органических молекул. Такие мутанты называются ауксотрофами (т. е. требующими дополнительного питания). При изучении организации бактериальных генов мы будем рассматривать ауксотрофные мутации только в качестве генетических маркеров. Более подробно они будут обсуждаться в главе 10. Фенотип ауксотрофных бактерий обозначают латинскими буквами, указывающими соединение, которое необходимо добавлять в среду для их нормального роста. Например, Met–, Thi– и Pur – обозначают, соответственно, мутантные штаммы, нуждающиеся в метионине, тиамине и пурине; соответствующие прототрофные фенотипы (дикий тип) обозначаются символами Met+, Тhi+ и Pur+. Мутации в самых различных генах могут давать одинаковые ауксотрофные фенотипы, и соответствующие генотипы обозначают теми же буквосочетаниями, что и фенотипы, но курсивом. Например, мутации metА и metB - это мутантные аллели генов дикого типа metА + и metB +, причем каждый мутант характеризуется фенотипом Met–. Как мы увидим далее, каждый из этих генов дикого типа необходим для биосинтеза метионина. Е. coli может использовать в качестве источника углерода многие органические соединения, более сложные чем глюкоза, поскольку обладает способностью превращать молекулы сложных Сахаров в молекулы
8. Бактериальный геном 229 глюкозы или других простых Сахаров, а также способностью расщеплять другие типы сложных молекул, например, аминокислот или жирных кислот. Такое расщепление сложных молекул называется катаболизмом. Мутации, затрагивающие катаболические функции, ограничивают типы молекул, которые могут служить источниками углерода для соответствующего мутанта. Например, мутант с фенотипом Lac ~ -не способен к росту в условиях, когда единственным источником углерода служит лактоза (молочный сахар), тогда как дикий тип Lac+ способен утилизировать лактозу. Фенотип Lac ~ может возникать в результате мутаций в генах lacZ + и lacY+, обусловливающих появление мутантных генотипов lacZ и lacY, соответственно. Заметим, что для того, чтобы определить, способен ли данный штамм к росту на определенной среде, надо знать, затрагивает ли соответствующая мутация анаболическую или катаболическую функцию. Например, мутант Met ~ нуждается для нормального роста в среде, обогащенной метионином, тогда как мутант Lac ~ не может расти на среде, содержащей в качестве источника углерода только лактозу и нуждается в другом источнике углерода. Оба типа мутаций служат удобными условно летальными генетическими маркерами. Температурочувствительные мутации широко используются и в генетике бактерий. Необходимые для нормального существования (существенные) гены, которые невозможно выявить посредством ауксотрофных мутаций, обычно могут быть идентифицированы с помощью температурочувствительных мутаций. Примерами жизненноважных функций могут служить функции, связанные с синтезом белков или нуклеиновых кислот из молекул-предшественников-аминокислот или нуклеотидов (подробное обсуждение мутаций, затрагивающих синтез ДНК, содержится в главе 13). Существуют также мутации бактерий, вызывающие устойчивость к определенным бактериофагам или антибиотикам. Первые из них обычно влияют на способность фага прикрепляться к бактерии-мутанту, поскольку у мутанта несколько изменены белки мембраны. Устойчивые (резистентные) мутанты легко отбираются при посеве клеток, подвергнутых действию какого-либо мутагена, непосредственно на среду, содержащую данный фаг или антибиотик: выросшая бактериальная колония и есть мутант. Мутации, вызывающие устойчивость к определенным антибиотикам, хорошо известны, поскольку они создают серьезную проблему для медицины и здравоохранения. Фенотип устойчивости к фагу Т1, обозначается символом T1R (фенотип чувствительности к фагу-XIs). Соответственно, устойчивость и чувствительность к антибиотику стрептомицину обозначают как StrR и StrS. Ген чувствительности к фагу Т1 обозначается как ton, (ton+ -синоним XIS; ген чувствительности к стрептомицину-sir (str+ -синоним strS). Мутантные штаммы легко получить при действии на бактерии дикого типа мутагенных факторов, например, рентгеновских или ультрафиолетовых лучей, или химических мутагенов. Обработанную культуру высевают на пермиссивную среду. Например, если нужно получить ауксотроф по определенной аминокислоте, то сперва подвергнутые действию мутагена бактерии высевают на среду, содержащую все 20 аминокислот. После появления колоний, их перепечатывают на чашку с минимальной средой, для того чтобы определить, какие из этих колоний ауксотрофные (рис. 8.1). Затем каждая из ауксотрофных колоний с пер-
230 Организация и передача генетического материала
миссивной среды «перепечатывается» на 20 чашек Петри с минимальным агаром, к которому добавлено по одной аминокислоте. При этом выясняется, присутствие какой именно аминокислоты необходимо для роста мутанта. Для отбора температурочувствительных мутантов обработанные мутагеном клетки высевают на чашку и инкубируют при 30°С. Выросшие колонии перепечатывают и чашку-реплику инкубируют при 42°С. Колонии, растущие при пермиссивной температуре 30°С, но не способные к росту при 42°С, образованы ts-мутантами. Подвергая полученные мутанты повторному (и последующему) воздействию мутагена и производя соответствующий отбор, можно получать штаммы, содержащие по нескольку мутаций. Дата добавления: 2015-12-16 | Просмотры: 1110 | Нарушение авторских прав |