|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Обзор результатов генетического анализа
В главах 5-8 мы видели, как генетический анализ позволяет определить общую организацию генетического материала у эукариотических и прокариотических организмов, и их вирусов. Комплементационный тест позволяет относить мутации к различным функциональным единицам. С помощью рекомбинационного анализа удается устанавливать взаимное расположение этих единиц и строить генетическую карту, представляющую собой модель хромосомной организации функциональных генетических единиц. Мы увидели, что генетические модели организации наследственного материала очень хорошо соответствуют реальной физической структуре ДНК. Кроме того, мы убедились в том, что генетический анализ может использоваться для изучения изменений тонкой структуры генов - изменений, затрагивающих отдельные пары нуклеотидов.
В последней главе первой части мы рассмотрим, как генетики используют генетические элементы прокариот для исследования тонкой структуры генетической организации прокариот и эукариот. Эти новые методы заложили основы «рекомбинантной революции в исследовании ДНК» и привели к появлению генетической инженерии. Изучив физическую и генетическую организацию генома, во второй части нашей книги мы вернемся к рассмотрению механизмов функционирования генов и к изучению того, каким образом гены определяют фенотип организма.
Литература
| Bukhari A.I., Shapiro J.A., Adhya S.L., eds., 1977. DNA Insertion Elements, Plasmids, and Episomes, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
Jacob F., Wollman E., 1961. Sexuality and the Genetics of Bacteria, Academic Press, New York.
Miller J. H., éd., 1972. Experiments in Molecular
| Genetics, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
Yanofsky C., Lennox E. (1959). Transduction and recombination study of linkage relationships among the genes controlling tryptophan synthesis in Escherichia coli, Virology, 8,
425-447.
|
Ключевые слова и понятия
| Анаболизм
Ауксотроф
Катаболизм
Инсерция (IS)
Конъюгация
Конъюгационная трубка
Конъюгационное картирование
Котрансдукция
| Мерозигота
Неселективный маркер
Общая трансдукция
Перепечатывание колоний
Плазмида
Прерванная конъюгация
Репликон
Селективный маркер
|
256 Организация и передача генетического материала
| Специфическая трансдукция
Трансдукция
Трансдукционное картирование
Транспозон
Трансформация
Фактор устойчивости к антибиотикам
| Эписома
Hfr-штамм
F-фактор
F'-штамм
F + -штамм
F – -штамм
|
Задачи
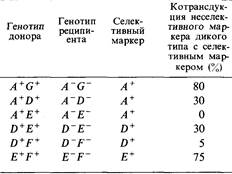
8.1. Последовательность передачи генов у различных Hfr-штаммов Е. coli различна; для нескольких штаммов она представлена ниже. Постройте на основе этих данных физическую карту бактериального генома.
 8.2. Для определения взаимного расположения генов А, В, С и D на генетической карте использовали метод прерванной конъюгации. Hfr-штамм leu + А +,В +,С +, D + Str передающий ген leu + в самом начале скрещивания, скрещивали со штаммом F – leu – А – ,В – ,С – ,D – StrR. Определите взаимное расположение генов А, В, С и D, используя гены StrR и leu + в качестве селективных маркеров (рис. 8.20).
8.2. Для определения взаимного расположения генов А, В, С и D на генетической карте использовали метод прерванной конъюгации. Hfr-штамм leu + А +,В +,С +, D + Str передающий ген leu + в самом начале скрещивания, скрещивали со штаммом F – leu – А – ,В – ,С – ,D – StrR. Определите взаимное расположение генов А, В, С и D, используя гены StrR и leu + в качестве селективных маркеров (рис. 8.20).
 Продолжительность конъюгации (мин)
Рис. 8.20. Графическое представление результатов скрещивания.
Продолжительность конъюгации (мин)
Рис. 8.20. Графическое представление результатов скрещивания.
| 8.3. Для определения взаимного расположения нескольких ауксотрофных мутаций Е. coli проводили эксперимент по прерываемому скрещиванию между прототрофным штаммом HfrStrS и штаммом F –, А – ,В – ,D –, StrR. В качестве селективных маркеров использовали устойчивость к стрептомицину и прототрофность по D. Определите расположение генов А и В относительно D, считая, что D + передается при скрещивании последним.
 8.4. Для определения взаимного расположения нескольких ауксотрофных мутаций Е. coli использовали данные по трансдукции фагом Р1. Проанализируйте приведенную таблицу и установите порядок следующих генов: А, В, С, D к F.
8.4. Для определения взаимного расположения нескольких ауксотрофных мутаций Е. coli использовали данные по трансдукции фагом Р1. Проанализируйте приведенную таблицу и установите порядок следующих генов: А, В, С, D к F.
|
8. Бактериальный геном 257
8.5. При скрещивании F ~ -штамма с различными Hfr-штаммами последовательность переноса генов различна. В таблице приводится порядок переноса различными Hfr-штаммами нескольких генов, расположенных неподалеку от точки начала (0). На основании этих данных постройте физическую карту бактериальной хромосомы.
 8.6. Для того, чтобы определить положение гена аrд 7 + на карте хромосомы Е. coli, прототрофный штамм StrSHfr4 (см. условие предыдущей задачи) скрещивали со штаммом F–,Met– Mtl– StrRarg7 –. После того как скрещивание прерывалось, образцы культуры высевали на чашки, содержавшие глюкозу в качестве источника углерода, стрептомицин и аргинин (или метионин). По приведенным в таблице данным определите расположение гена arg7 +.
8.6. Для того, чтобы определить положение гена аrд 7 + на карте хромосомы Е. coli, прототрофный штамм StrSHfr4 (см. условие предыдущей задачи) скрещивали со штаммом F–,Met– Mtl– StrRarg7 –. После того как скрещивание прерывалось, образцы культуры высевали на чашки, содержавшие глюкозу в качестве источника углерода, стрептомицин и аргинин (или метионин). По приведенным в таблице данным определите расположение гена arg7 +.
 8.7. Студент-генетик получил задание определить положение на карте ауксотрофной мутации, которую он на основа-
8.7. Студент-генетик получил задание определить положение на карте ауксотрофной мутации, которую он на основа-
| нии данных по трансдукции считает тесно сцепленной с геном his. Он ставит конъюгационное скрещивание штамма Hfr АВ313 StrSAde – TIS (см. условие задачи 8.51 с ауксотрофным штаммом F–  . После прерывания конъюгации образцы суспензии он высевает на среду, содержащую глюкозу и стрептомицин. Рекомбинантов обнаруживается очень мало, и их недостаточно для того, чтобы установить тесное сцепление. Со своими трудностями он обращается к преподавателю. Тот советует ему исключить из среды стрептомицин и повторить эксперимент. Студент пользуется советом и получает надежные данные, показывающие, что ген, положение которого он должен определить, попадает в F ~ -клетки за две минуты до гена his. Объясните, что было неправильно в первом эксперименте, и почему во втором случае эксперимент удался. Предложите другую схему эксперимента.
8.8. Изобразите схематически рекомбинанты, которые могут быть выделены из нового F'-штамма, в котором F'-элемент содержит гены thr + leu –, a бактериальная хромосома - гены thr – leu +. Обозначьте положение полового фактора в каждом рекомбинанте.
8.9. Предложите два способа определения того, принадлежит ли отобранная после скрещивания Hfr и F– -штаммов рекомбинантная колония типу F' или F–.
8.10. Установлено, что определенный штамм Е. coli, требующий для роста метионин, содержит в гене met элемент IS1, инактивирующий этот ген и вызывающий потребность в метионине. Из этого штамма выделено множество спонтанных мутантов с делециями, затрагивающими смежный по отношению к гену met оперон ilv. Носители этих делеций, кроме метионина, нуждаются также в изолейцине и валине. Каждый из штаммов с одной из делеций используется в качестве донорного для трансдукции фагом Р1 маркеров ilv + в реципиентные штаммы ilv –. Используются реципиентные штаммы ilv А –, ilvB –, ilvC –и ilvD –. В приводимой таблице указано присутствие (+) или отсутствие (—) рекомбинантов ilv + в каждом . После прерывания конъюгации образцы суспензии он высевает на среду, содержащую глюкозу и стрептомицин. Рекомбинантов обнаруживается очень мало, и их недостаточно для того, чтобы установить тесное сцепление. Со своими трудностями он обращается к преподавателю. Тот советует ему исключить из среды стрептомицин и повторить эксперимент. Студент пользуется советом и получает надежные данные, показывающие, что ген, положение которого он должен определить, попадает в F ~ -клетки за две минуты до гена his. Объясните, что было неправильно в первом эксперименте, и почему во втором случае эксперимент удался. Предложите другую схему эксперимента.
8.8. Изобразите схематически рекомбинанты, которые могут быть выделены из нового F'-штамма, в котором F'-элемент содержит гены thr + leu –, a бактериальная хромосома - гены thr – leu +. Обозначьте положение полового фактора в каждом рекомбинанте.
8.9. Предложите два способа определения того, принадлежит ли отобранная после скрещивания Hfr и F– -штаммов рекомбинантная колония типу F' или F–.
8.10. Установлено, что определенный штамм Е. coli, требующий для роста метионин, содержит в гене met элемент IS1, инактивирующий этот ген и вызывающий потребность в метионине. Из этого штамма выделено множество спонтанных мутантов с делециями, затрагивающими смежный по отношению к гену met оперон ilv. Носители этих делеций, кроме метионина, нуждаются также в изолейцине и валине. Каждый из штаммов с одной из делеций используется в качестве донорного для трансдукции фагом Р1 маркеров ilv + в реципиентные штаммы ilv –. Используются реципиентные штаммы ilv А –, ilvB –, ilvC –и ilvD –. В приводимой таблице указано присутствие (+) или отсутствие (—) рекомбинантов ilv + в каждом
|
258 Организация и передача генетического материала
опыте. Определите расположение мутаций ilv –друг относительно друга и относительно гена met.
 8.11. Мутанты thr и leu у Е. coli - ауксотрофы, требующие для роста треонин и лейцин, соответственно. Мутация аra3 обусловливает неспособность клеток использовать арабинозу в качестве источника углерода. В приводимой таблице указаны частоты котрансдукции этих генов фагом Р1. Какая селективная среда использовалась в каждом случае и какова последовательность генов?
8.11. Мутанты thr и leu у Е. coli - ауксотрофы, требующие для роста треонин и лейцин, соответственно. Мутация аra3 обусловливает неспособность клеток использовать арабинозу в качестве источника углерода. В приводимой таблице указаны частоты котрансдукции этих генов фагом Р1. Какая селективная среда использовалась в каждом случае и какова последовательность генов?
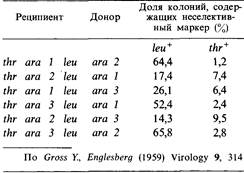
 8.12. Мутации ara1 и ara 2 очень тесно сцеплены с агаЗ. Все они делают клетки Е. coli неспособными к использованию арабинозы в качестве единственного источника углерода. Для того, чтобы установить взаимное расположение этих мутаций, проводили реципрокные трансдукционные скрещивания, результаты которых представлены в таблице. Во всех случаях отбирали рекомбинанты Ara + и среди них определяли долю носителей неселективных маркеров leu + и thr +. Каково взаимное расположение этих маркеров?
8.12. Мутации ara1 и ara 2 очень тесно сцеплены с агаЗ. Все они делают клетки Е. coli неспособными к использованию арабинозы в качестве единственного источника углерода. Для того, чтобы установить взаимное расположение этих мутаций, проводили реципрокные трансдукционные скрещивания, результаты которых представлены в таблице. Во всех случаях отбирали рекомбинанты Ara + и среди них определяли долю носителей неселективных маркеров leu + и thr +. Каково взаимное расположение этих маркеров?
|  8.13. Когда в качестве реципиентных используются клетки агаЗ, а донорными клетками для фага Р1 служат ara1, ara 2 или клетки дикого типа, и отбираются трансдуктанты Ara +, то на селективной среде наряду с крупными колониями появляются крошечные колонии, неразличимые невооруженным глазом. Сформулируйте гипотезу об их происхождении.
8.14. Клетки некоторых штаммов Salmonella paratyphi подвижны, поскольку обладают жгутиками; клетки других штаммов лишены жгутиков и неспособны к самостоятельному движению. Фаг, осуществляющий общую трансдукцию у Salmonella (P22), выращивают на подвижных клетках и инфицируют им неподвижные клетки. Когда инфицированные клетки неподвижного штамма высевают на поверхность столбика из мягкого агара, то после инкубации можно увидеть тянущиеся вглубь агара от поверхности цепочки мелких колоний (рис. 8.21).
Рис. 8.21. Бактериальные колонии Salmonella paratyphi в пробирке с мягким агаром. [ Leaerberg J. (1956). Genetics, 41, 845.]
8.13. Когда в качестве реципиентных используются клетки агаЗ, а донорными клетками для фага Р1 служат ara1, ara 2 или клетки дикого типа, и отбираются трансдуктанты Ara +, то на селективной среде наряду с крупными колониями появляются крошечные колонии, неразличимые невооруженным глазом. Сформулируйте гипотезу об их происхождении.
8.14. Клетки некоторых штаммов Salmonella paratyphi подвижны, поскольку обладают жгутиками; клетки других штаммов лишены жгутиков и неспособны к самостоятельному движению. Фаг, осуществляющий общую трансдукцию у Salmonella (P22), выращивают на подвижных клетках и инфицируют им неподвижные клетки. Когда инфицированные клетки неподвижного штамма высевают на поверхность столбика из мягкого агара, то после инкубации можно увидеть тянущиеся вглубь агара от поверхности цепочки мелких колоний (рис. 8.21).
Рис. 8.21. Бактериальные колонии Salmonella paratyphi в пробирке с мягким агаром. [ Leaerberg J. (1956). Genetics, 41, 845.]

|
8. Бактериальный геном 259
В культурах, выделенных из этих колоний, все клетки оказываются неподвижными. Объясните.
8.15. При некоторых скрещиваниях с участием лизогенных штаммов профаги типа λ могут использоваться в качестве бактериального генетического маркера. В таблице представлены результаты скрещивания между нелизогенным [(1у)–]
 штаммом HfrH,Str и штаммами F Thr– Leu– Gal– StrR, лизогенными по одному из нескольких различных профагов, например, F–,Thr– Leu– Gal– StrR (λ). Отбираются рекомбинанты Thr +
штаммом HfrH,Str и штаммами F Thr– Leu– Gal– StrR, лизогенными по одному из нескольких различных профагов, например, F–,Thr– Leu– Gal– StrR (λ). Отбираются рекомбинанты Thr +
| Leu + StrR или Gal + StrR и среди них определяют долю (ly–)Hfr. О наличии маркера ly– судят по чувствительности к соответствующему фагу, другими словами, утрата профага рекомбинантом интерпретируется как наличие неселективного маркера. На основании этих данных определите взаимное расположение профагов на физической карте Е. coli и их положение относительно Thr + Leu + и Gal +.
8.16. При скрещивании штамма Hfr, лизогенного по фагу λ, с нелизогенным F– -штаммом может иметь место феномен, называемый зиготической индукцией. Что это такое можно понять, сравнивая рекомбинанты, получаемые в реципрокных скрещиваниях между содержащими профаги штаммами HfrHStr и F–,Thr– Leu–ArBTlRLac–gal–StrB, результаты которых приведены в таблице. Основываясь на представленных в таблице данных о частоте возникновения рекомбинантов при зиготической индукции, постройте гипотезу, объясняющую этот феномен, и предложите постановку эксперимента для проверки вашей гипотезы.
| 
|
Дата добавления: 2015-12-16 | Просмотры: 913 | Нарушение авторских прав
|