|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
F-фактор: генетический элемент, определяющий пол бактерийF-фактор обладает поразительной способностью определять пол, казалось бы, бесполых бактерий. Его существование было обнаружено генетиками, когда они пытались определить, происходят ли скрещивания
232 Организация и передача генетического материала
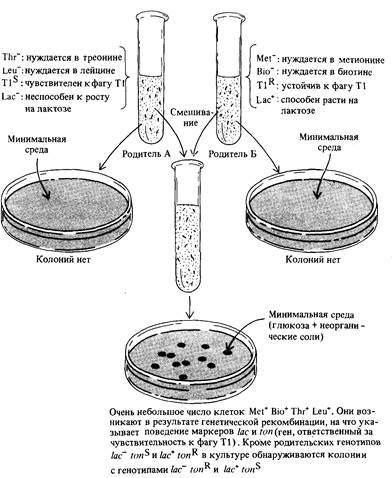
между различными линиями Е. coli. Для этого исследовали возможность генетической рекомбинации между различными мутантными штаммами. Множественные ауксотрофы бактерий, полученные посредством нескольких последовательных этапов мутагенеза и селекции, смешивали и смесь высевали на минимальную среду. Появление колоний должно было свидетельствовать о возникновении рекомбинантов дикого типа, как это изображено на рис. 8.2. Использование множественных ауксотрофов гарантирует, что колонии дикого типа, способные к росту на минимальной среде, возникают именно в результате рекомбинации, а не вследствие обратных мутаций. Если штамм, ауксотрофный по одной аминокислоте, ревертирует к дикому типу с частотой 1·106, то появление колоний дикого типа на минимальной среде можно расценить и как результат реверсии, и как результат рекомбинации. Если же мутант ауксотрофен по двум аминокислотам, то частота реверсии к дикому типу должна составлять 106 · 106 = = 1012. Однако, при использовании таких двойных ауксотрофов в эксперименте колонии дикого типа на минимальной среде образовы-
8. Бактериальный геном 233
вались с частотой примерно 1·105, т.е. много чаще, чем этого можно было бы ожидать, основываясь на предположении об обратных мутациях. Эти эксперименты обнаружили существование у Е. coli двух половых типов, обозначенных символами F + и F –. Затем было установлено, что клетки типа F + содержат F-фактор, или эписому, тогда как в клетках F ~ -типа эта эписома отсутствует. Дальнейшие исследования бактерий двух типов привели к поразительному и совершенно неожиданному открытию: оказалось, что Fфактор инфекционен. Когда обладавшие F-фактором и чувствительные к стрептомицину клетки (StrS, F +) смешивали со стрептомицинустойчивыми бактериями (StrR, F –), и эту смесь высевали на агар, содержащий стрептомицин, то колонии образовывали, естественно, лишь StгR-клетки, т. е. клетки, несущие аллель sirR. Большинство из этих колоний оказалось F + -, а не F – -типа. F-фактор содержит множество генов, сообщающих ему инфекционность. Некоторые из этих генов кодируют белки пилей, структур типа трубочек, расположенных на поверхности F + -клеток (рис. 8.3). F-пили соединяются с соответствующими рецепторами на поверхности F ~ -клеток, что приводит к образованию цитоплазма тического мостика между двумя клетками. В процессе роста F+клеток F-фактор реплицируется по тета-типу, так же как и бактериальная хромосома. Однако, когда между F + - и F – -клетками возникает цитоплазматический мостик, F-фактор переходит к реплика-
234 Организация и передача генетического материала
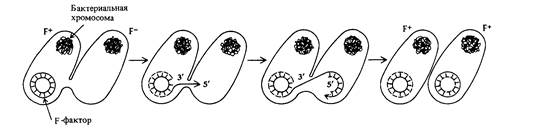
ции по сигма-типу (см. гл. 4). 5'-РО4 одноцепочечный конец реплицирующегося F-фактора проникает в F – -клетку, и там синтезируется комплементарная цепь. Репликация ДНК F-фактора приводит к переносу копии F-фактора в F – -клетки, превращая их в F + -клетки (рис. 8.4). Как уже упоминалось ранее, F-фактор представляет собой эписому, которая может либо существовать самостоятельно, либо встраиваться в репликон бактерии. В встроенном состоянии F-фактор может переносить бактериальную хромосому в F – -клетки. Частота возникновения рекомбинантов дикого типа для генов бактериальной хромосомы при скрещивании F + - и F – -штаммов очень низка (порядка одной клетки на 105 родительских клеток), поскольку лишь небольшое число клеток F+культуры участвует в образовании рекомбинантов, хотя частота инфицирования F-фактором довольно высока. Однако, из F + -культуры можно выделить штаммы, при скрещивании которых с F – -клетками рекомбинанты образуются гораздо чаще (частота рекомбинации > 10–2). Эти штаммы обозначаются символом Hfr (от англ, high frequency recombination -высокая частота рекомбинации). В них F-фактор в свободном, автономном состоянии отсутствует, он встроен в бактериальную хромосому. Когда клетки Hfr вступают в контакт с клетками F –, между ними образуется цитоплазматический мостик, называемый конъюгационной трубкой, и интегрированный F-фактор инициирует репликацию бактериальной хромосомы по типу катящегося кольца с того сайта, в который он встроен. Эта репликация приводит к переносу бактериальной хромосомы в F – -клетку (рис. 8.5). Когда копия бактериальной хромосомы клетки Hfr попадает в клетку F –, становится возможной рекомбинация между генетическими маркерами, содержащимися в хромосоме клетки Hfr, и генами, содержащимися в хромосоме F– -клетки (рис. 8.5). При конъюгации клеток Hfr и F– часто происходит разрушение соединяющей их конъюгационной трубки и, соответственно, обрыв хромосомы Hfr. В результате в F–клетку целая Hfr-хромосома попадает довольно редко. После того, как конъюгация инициирована, возникновение рекомбинантов дикого типа определяется исключительно жизнеспособностью F –-родителя. К такому выводу можно прийти при анализе результатов реципрокных скрещиваний, выполняемых на минимальной среде, содержащей стрептомицин.
8. Бактериальный геном 235
Скрещивание 1: Hfr, Thr + Leu + Strs x F –, Thr – Leu – StrR Результат: колонии образуют лишь рекомбинанты Thr+ Leu+ StrR. Скрещивание 2: Hfr, Thr– Leu– StrRx F–, Thr+ Leu+ Str5. Результат: колоний нет. Первое скрещивание показывает, что применение стрептомицина устраняет колонии, которые в противном случае образовались бы из клеток родителя Hfr. Формировать колонии могут только рекомбинанты дикого типа, поскольку родители F – -типа не могут расти на минимальной среде в отсутствие треонина и лейцина. Второе скрещивание показывает, что для образования рекомбинантов необходима жиз-
236 Организация и передача генетического материала неспособность родителя F ~, а не Hfr. Рекомбинантные колонии StrR не образуются, поскольку ген sfrR локализован слишком далеко от сайта, с которого начинается передача хромосомы и потому почти никогда не попадает в F – -клетку. То, что поступление бактериальных генов из Hfr в F – -клетку происходит последовательно, всегда начиная с сайта интеграции F-фактора в бактериальную хромосому, дает возможность картировать гены по порядку их попадания в F – -клетку (подробнее мы обсудим это в следующем разделе). Как показано на рис. 8.5, проникновение хромосомы в F – -клетку всегда начинается с нуклеотидной последовательности интегрированного F-фактора. При этом в F – -клетку поступает не весь Fфактор. Для того чтобы в клетку попала оставшаяся его часть, необходимо, чтобы в F – перешла вся Hfr-хромосома. Это случается очень редко из-за спонтанных разрывов конъюгационной трубки, и поэтому большинство отбираемых рекомбинантов остаются принадлежащими F – -типу. Дата добавления: 2015-12-16 | Просмотры: 1127 | Нарушение авторских прав |