|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
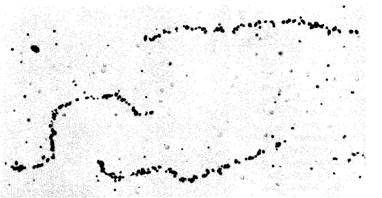
Организация генома фагов Т2 и Т4Крупные фаги Т2 и Т4 близкородственны. Они обладают идентичной организацией генома, и большинство генов у них общие. Радиоавтография хромосомы фага Т4 (рис. 7.13) свидетельствует о линейности молекулы ДНК. Ее мол. масса равна 120-106 дальтон, а длина составляет 182 000 пары нуклеотидов. Фаг Т4 был предметом интенсивных генети-
214 Организация и передача генетического материала
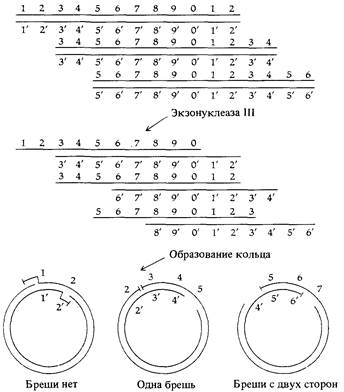
ческих исследований (вспомните rII -мутанты из гл. 6), и для него известны мутации во многих цистронах. Анализ рекомбинации с использованием трех- и четырехфакторных скрещиваний показал, что генетическая карта фага Т4 имеет кольцевую форму (рис. 7.14). Противоречие между линейностью молекулы ДНК фага и кольцевой формой его генетической карты удалось разрешить в результате генетических и физических экспериментов, показавших, что выделенные из фага Т2 линейные молекулы ДНК содержат на обоих концах участки с одинаковыми последовательностями нуклеотидов (концевую избыточность), а порядок генов в молекуле допускает циклические перестановки. Физические данные о концевой избыточности (дуплицированности) последовательностей оснований получены из экспериментов, в которых ДНК Т2 подвергалась действию экзонуклеазы III. Этот фермент последовательно отщепляет нуклеотиды с 3-ОН-концов цепей ДНК, в результате чего на концах двухцепочечной молекулы образуются одноцепочечные участки с 5'-РО4-концами (рис. 7.15). Инкубация этих подвергнутых действию фермента молекул в условиях, допускающих установление водородных связей между комплементарными одноцепочечными последовательно-
7. Геном вируса 215
стями, приводит к образованию кольцевых молекул (рис. 7.16). Замыкание в кольцо возможно лишь в том случае, если двухцепочечная молекула ДНК фага Т2 содержит идентичные последовательности оснований на обоих концах, как это схематически изображено на рис. 7.15. Прежде чем молекулы становятся способны образовывать кольца, фермент должен отщепить около 2% ДНК Т2. Это означает, что протяженность дуплицированных (избыточных) концов молекулы составляет около 1% длины генома. Генетические данные о концевой избыточности геномов фагов Т2 и Т4 проистекают из существования гетерозиготных фаговых частиц. Фаговые гетерозиготы возникают в результате рекомбинации между фагами с различными генотипами. Они легко идентифицируются, поскольку каждый такой фаг образует негативную колонию, содержащую фаги обоих генотипов (см. рис. 6.1). Геном единичного фага может быть
216 Организация и передача генетического материала
гетерозиготным лишь по тесно сцепленным генам. В популяции фагов, однако, встречаются гетерозиготы по всем генам, безотносительно к их положению на карте. Кроме того, размер участка, по которому фаг избыточен и гетерозиготен, увеличивается у мутантов, несущих делеции, в связи с особым механизмом упаковки, который мы вкратце рассмотрим ниже. (В отличие от фага λ у фагов Т2 и Т4 делеции не уменьшают количество ДНК в головке фага.) Из этих наблюдений можно сделать лишь один логический вывод: не существует единственного уникального порядка генов вдоль всей молекулы ДНК, другими словами, у конкретных фагов в популяции избыточным может быть любой ген.
7. Геном вируса 217
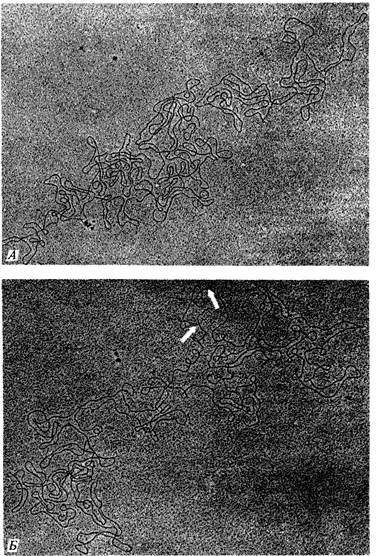
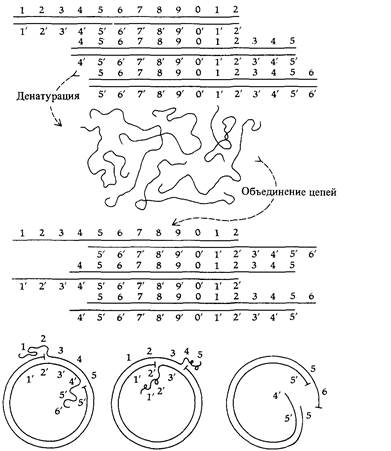
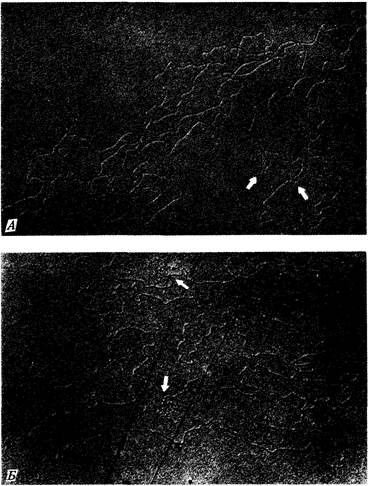
Следующий эксперимент подтверждает это поразительное заключение. Препарат ДНК фага Т2 денатурировали нагреванием, для того чтобы разделить комплементарные цепи каждой молекулы. Смесь одиночных цепей выдерживали затем в условиях, допускающих восстановление водородных связей между комплементарными последовательностями оснований. Большинство одиночных цепей в смеси оказалось способно восстановить двухцепочечную структуру с партнером, исходно принадлежавшим другой нативной молекуле ДНК (рис. 7.17). При электронно-микроскопическом анализе такой смеси обнаруживается много кольцевых двухцепочечных молекул (рис. 7.18). Образование двухцепочечных структур возможно лишь в том случае, если исходный препарат ДНК содержит такую популяцию молекул, в которой последовательность генов в любой молекуле можно получить посредством циклической перестановки (пермутации) генов в любой другой молекуле. Именно циклические перестановки и концевая избыточность индивидуальных молекул ДНК фагов Т2 и Т4 обусловливают кольцевую структуру их генетической карты, отражая отношения сцепления между генами в популяции индивидуальных молекул.
218 Организация и передача генетического материала
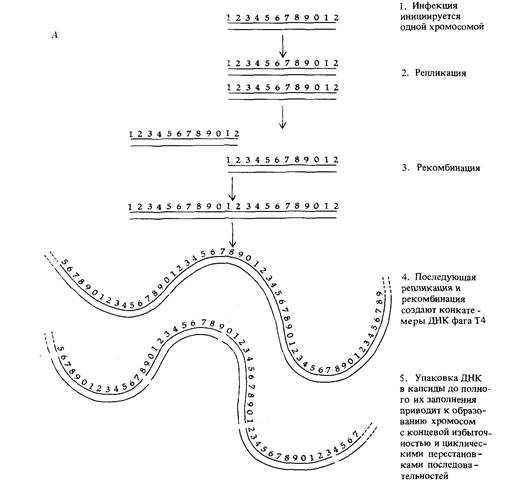
Специальный способ репликации ДНК и упаковки дочерних молекул ДНК в головки фагов Т2 и Т4 обеспечивает возможность получения от единичных фаговых частиц потомства с циклической перестановкой и концевой избыточностью. На ранних стадиях инфекции линейная родительская молекула ДНК претерпевает несколько последовательных репликаций, образуя такие же линейные дочерние молекулы, содержащие весь геном фага плюс концевую избыточность. Затем в результате рекомбинации между избыточными концами дочерних молекул образуются конкатемеры (тандемно повторяющиеся последовательности геномов фага (рис. 7.19), которые затем реплицируются и рекомбинируют, образуя еще более длинные конкатемеры. На заключительной стадии инфекции молекулы ДНК начинают упаковываться в головки (капсиды) фагов (вспомните рис. 7.2). Размер молекулы ДНК, помещае-
7. Геном вируса 219
мой в капсид каждого фага, определяется размером самого капсида. Капсид вмещает чуть больше ДНК, чем ее содержится в геноме фага Т4, как раз настолько, чтобы осталось место для избыточных участков на концах генома. Такой механизм упаковки ДНК посредством отрезания от длинного конкатемера кусков, по длине несколько превышающих протяженность генома фага, приводит к циклической перестановке (пермутации) последовательности генов в отдельных дочерних фагах,
220 Организация и передача генетического материала
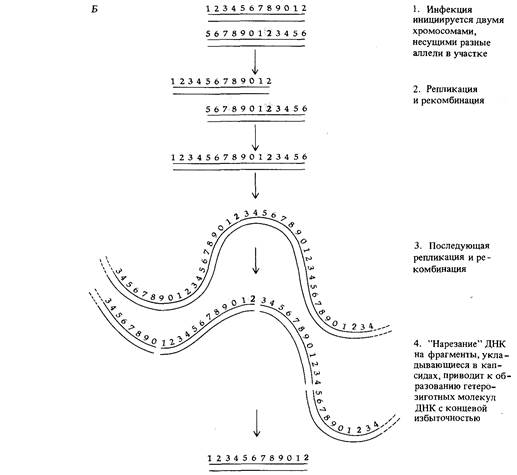
как это схематически показано на рис. 7.19. Тот же самый механизм инкапсулирования геномов фага Т4 приводит к возникновению гетерозигот с избыточными концами в двукратно инфицированных клетках (рис. 7.19). Такой тип организации генома встречается не только у бактериофагов. Он обнаружен также, например, у так называемого «вируса 3 лягушки» - вируса, способного инфицировать клетки многих видов позвоночных, в том числе и человека, при температуре ниже 32°С.
7. Геном вируса 221
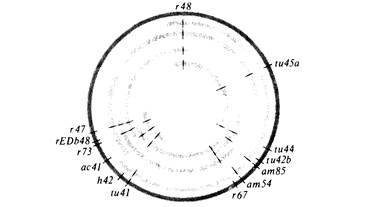
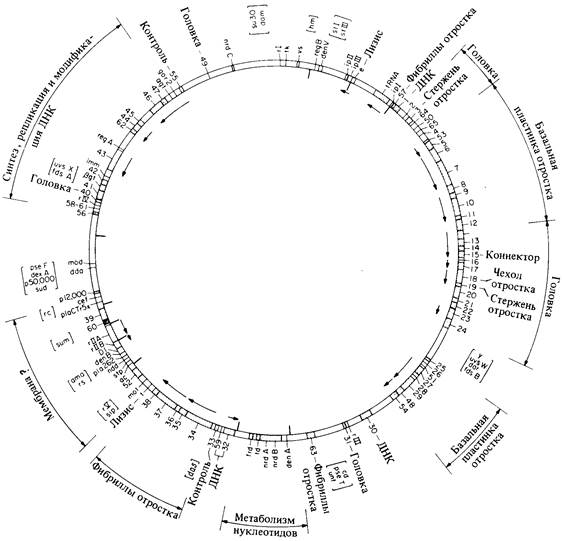
На генетической карте фага Т4 (рис. 7.20) видно, что вместе сгруппированы гены, ответственные за родственные физиологические функции. Такая же организация генома уже встречалась нам у фага λ. Этот тип организации функционирования генома играет важную роль в ее регуляции (гл. 15).
222 Организация и передача генетического материала Литература
Дата добавления: 2015-12-16 | Просмотры: 1194 | Нарушение авторских прав |