|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология
|
Ключевые слова и понятия. Белок-репрессор Генетическая карта Гетеродуплекс Двухфакторное скрещивание Индукция профага Конкатемер Концевая избыточность
| Белок-репрессор
Генетическая карта
Гетеродуплекс
Двухфакторное скрещивание
Индукция профага
Конкатемер
Концевая избыточность последовательности ДНК
Лизогенизация
Липкие концы
Литический путь
Морфогенез фага Т4
Мутации amber, ochre, opal
| Мутации clear фага λ
Неселективный маркер
Низкая отрицательная интерференция
Одиночный цикл развития фага
Профаг
Селективный маркер
Супрессорчувствительные мутации
Температурочувствительные мутации
Трехфакторное скрещивание
Физическая карта
Циклическая перестановка (пермутация)
последовательности ДНК
|
Задачи
7.1. В таблице представлены результаты комплементационного теста между различными температурочувствительными фаговыми мутантами; «+» означает присутствие негативных колоний при непермиссивной температуре. Распределите мутации по группам комплементации.

| 7.2, Определите условия эксперимента при постановке теста на комплементацию между следующими парами условно летальных мутаций фага: a) amber, amber;
б) amber, ochre;
в) теплочувствительная, теплочувствительная;
г) холодочувствительная, холодочувствительная;
д) amber и теплочувствительная;
е) теплочувствительная, холодочувствительная.
7.3. Перечислите пермиссивные и непермиссивные условия для каждой из мутаций и селективные маркеры при постановке трехфакторных скрещиваний в следующих комбинациях мутантов:
|
7. Геном вируса 223
a) amber, amber, температурочувствительная;
б) opal, температурочувствительная, температурочувствительная;
в) ochre, ochre, amber;
г) морфология негативных колоний, amber, температурочувствительная;
д) ochre, opal, морфология негативных колоний;
е) специфичность к хозяину, специфичность к хозяину, размер негативных колоний.
7.4. Проведены двухфакторные скрещивания между четырьмя температурочувствительными мутантами. Цифры в таблице означают долю рекомбинантов дикого типа в потомстве при культивировании при 40°С. Определите последовательность генов на генетической карте.
 7.5. Проведены скрещивания температурочувствительных мутантов мелкого фага с кольцевым геномом. Проанализируйте таблицу и постройте карту хромосомы фага.
7.5. Проведены скрещивания температурочувствительных мутантов мелкого фага с кольцевым геномом. Проанализируйте таблицу и постройте карту хромосомы фага.
 7.6. На мутантах мелкого фага были поставлены два указанных ниже трехфакторных скрещивания. Считайте, что селективные маркеры всегда теснее сцеплены по сравнению с неселективными.
7.6. На мутантах мелкого фага были поставлены два указанных ниже трехфакторных скрещивания. Считайте, что селективные маркеры всегда теснее сцеплены по сравнению с неселективными.
| Постройте карту последовательности мутантных генов.
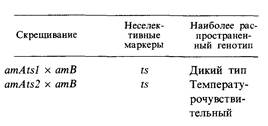 7.7. Были поставлены следующие трехфакторные скрещивания. Принимая предположение, которое было сделано в условии задачи 7.6, определите последовательность генов на генетической карте.
7.7. Были поставлены следующие трехфакторные скрещивания. Принимая предположение, которое было сделано в условии задачи 7.6, определите последовательность генов на генетической карте.
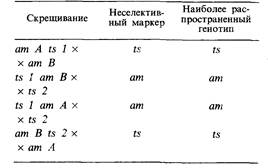
 7.8. На мутантах мелкого фага были поставлены следующие трехфакторные скрещивания. Принимая предположения, сделанные в условиях двух предыдущих задач, постройте карту последовательности генов.
7.8. На мутантах мелкого фага были поставлены следующие трехфакторные скрещивания. Принимая предположения, сделанные в условиях двух предыдущих задач, постройте карту последовательности генов.
|
224 Организация и передача генетического материала
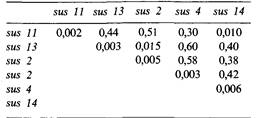
7.9. Для изучения комплементации между различными мутациями у фагов часто определяют разницу в их урожае в пермиссивных и непермиссивных клетках при смешанной инфекции. Проанализируйте таблицу и распределите мутации по комплементационным группам.
 7.10, ф105-это умеренный фаг; его хозяином является бактерия Bacillus subtilis. Фаговые частицы дикого типа образуют мутные негативные колонии при температуре как 30, так и 40°С. В двухфакторных скрещиваниях было картировано несколько температурочувствительных мутаций. Указанные в таблице цифры соответствуют доле (%) рекомбинантов дикого типа в соответствующих скрещиваниях, проведенных при температуре 40°С. Постройте по этим данным генетическую карту.
7.10, ф105-это умеренный фаг; его хозяином является бактерия Bacillus subtilis. Фаговые частицы дикого типа образуют мутные негативные колонии при температуре как 30, так и 40°С. В двухфакторных скрещиваниях было картировано несколько температурочувствительных мутаций. Указанные в таблице цифры соответствуют доле (%) рекомбинантов дикого типа в соответствующих скрещиваниях, проведенных при температуре 40°С. Постройте по этим данным генетическую карту.
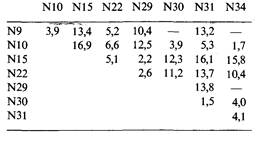 Мутант cc1 образует при обеих температурах прозрачные негативные колонии, а мутант с4 дает мутные негативные колонии при 30°С и прозрачные при 40°С. По представленным ниже данным постройте генетическую карту фага φ 105, включающую уже построенную ее часть.
Мутант cc1 образует при обеих температурах прозрачные негативные колонии, а мутант с4 дает мутные негативные колонии при 30°С и прозрачные при 40°С. По представленным ниже данным постройте генетическую карту фага φ 105, включающую уже построенную ее часть.
| 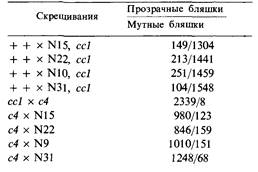 По Rutberg L (1969). J. Virol., 3, 38.
7.11. Проводили приведенные ниже трехфакторные скрещивания и каждый раз отмечали преобладающий генотип по неселективному маркеру. Во всех скрещиваниях селективные маркеры были теснее сцеплены друг с другом, чем каждый из них с неселективным (сцепленность предварительно определяли в двухфакторных скрещиваниях). Определите последовательность мутантных генов на генетической карте.
По Rutberg L (1969). J. Virol., 3, 38.
7.11. Проводили приведенные ниже трехфакторные скрещивания и каждый раз отмечали преобладающий генотип по неселективному маркеру. Во всех скрещиваниях селективные маркеры были теснее сцеплены друг с другом, чем каждый из них с неселективным (сцепленность предварительно определяли в двухфакторных скрещиваниях). Определите последовательность мутантных генов на генетической карте.
 7.12. Клетки Е. coli, лизогенные по фагу λ, устойчивы к нему и не могут быть инфицированы этим же фагом λ. Однако возможно одновременное инфицирование чувствительных клеток двумя фагами λ с различными генотипами; получаемые при этом клетки называются
7.12. Клетки Е. coli, лизогенные по фагу λ, устойчивы к нему и не могут быть инфицированы этим же фагом λ. Однако возможно одновременное инфицирование чувствительных клеток двумя фагами λ с различными генотипами; получаемые при этом клетки называются
|
7. Геном вируса 225
| двойными лизогенами и содержат в генотипе оба профага. Двойная инфекция фагами λ и λcI приводит к образованию лизогенов, содержащих оба профага, тогда как фаг λсI сам по себе никогда не лизогенизирует клетку даже в присутствии «хелпера» λсI+ во время инфекции. Опишите схематически возможную последовательность событий, приводящую к такой двойной лизогении. Что вы можете сказать относительно продукта гена сI+?
| 7.13. Первые генетические исследования фага λ относились к мутациям, изменяющим размер или морфологию негативных колоний. Вот некоторые из этих мутаций: s (small, мелкие бляшки), с (clear, прозрачные бляшки, а не мутные, впоследствии обозначенные символом cI), co1 и со2 (cocarde) прозрачные; в настоящее время для них приняты обозначения cIII и cII соответственно), mi (minute, очень мелкие бляшки, меньшие, чем у мутанта s). В таблице приведены данные по некоторым трехфакторным скрещиваниям этих мутантов. Постройте генетическую карту с указанием расстояний между соседними генами.
|

| |
| | По Kaiser A. (1957). Virology, 3, 42.
| |
7.14. Фаги λcΙΙ и λcΙΙΙ могут лизогенизировать клетки Е. coli в присутствии фагов хелперов λ + или λcI. Зная ответы на две предыдущие задачи, что вы можете сказать относительно cI, cII и cIII?
7.15. Все умеренные фаги лямбдоидного семейства обладают «липкими» концами. Большинство из них способно встраиваться в различные участки хромосомы хозяина и обладают иммунными свойствами, перечисленными ниже в таблице, где «+» означает успешное

| инфицирование, а «—» устойчивость к инфекции.
При смешанной инфекции некоторые лямбдоидные фаги могут обмениваться генами друг с другом. В прилагаемой таблице указывается присутствие рекомбинантов дикого типа с λ-иммунностью в потомстве от скрещивания различных мутантов фага λ с фагами дикого типа 434, 21 и 82. Какой из этих фагов наиболее близок фагу λ? Что вы можете сказать относительно генетического контро-

|
226 Организация и передача генетического материала
| ля иммунности и лизогенизации, судя по этой таблице и по трем предыдущим задачам?
7.16. В результате рекомбинации между фагом λ и фагом 434 иногда образуется гибридный фаг, в котором хромосома фага λ обладает иммунными свойствами фага 434. Такой рекомбинант обозначается символом λimm434; он спо-
| собен к росту на хозяине с профагом λ в геноме, но не на хозяине с профагом 434. Ген су локализован между геном О и участком, определяющим иммунные свойства. Для того чтобы определить порядок мутаций 2001 и 42 в гене су, были поставлены два скрещивания. Потомство высевали на Е. coli (λimm434Oam29). Каков порядок мутаций?
|
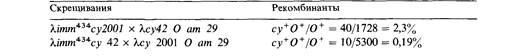
| | По Jones M.O., Herskowitz I. (1978). Virology, 88, 199.
|
7.17. Мутация фага λcαn1 делает возможной лизогенизацию мутантами λcΙΙΙ. Другими словами, λcIIIcαn1 образует мутные негативные колонии. λcαn+ можно отличить от λcαn1 при посеве на клетки Е. coli штамма WA8067: негативные колонии λcαn+ чуть более прозрачны, чем у λcan1. На основании приводимых ниже результатов скрещиваний определите положение сап1 относительно мутаций су, картированных в предыдущей задаче. Потомство высевали на Е. coli Su – (λimm4340am29).
 По Jones M. 0., Herskowitz I. (1978). Virology, 88, 199.
7.18. Прерывание скрещивания путем искусственного лизиса клетки на той стадии, когда в ней содержится всего около десятка фаговых частиц, служит одним из способов демонстрации кольцевой структуры генетической карты фага Т4. Если лизис проходит нормально, каждая
По Jones M. 0., Herskowitz I. (1978). Virology, 88, 199.
7.18. Прерывание скрещивания путем искусственного лизиса клетки на той стадии, когда в ней содержится всего около десятка фаговых частиц, служит одним из способов демонстрации кольцевой структуры генетической карты фага Т4. Если лизис проходит нормально, каждая
| клетка содержит около 200 фагов-потомков, и уже невозможно наблюдать сцепление между сравнительно удаленными маркерами, а именно это необходимо для доказательства кольцевой структуры. Исходя из содержащегося в тексте главы обсуждения динамики скрещивания фагов, сформулируйте гипотезу, объясняющую эти наблюдения. Как должно было бы влиять на наблюдаемые частоты рекомбинации прерывание нормального лизиса клеток в соответствии с вашей гипотезой? Лизис клеток не происходит, когда оба родительских фага мутантны по соответствующему гену (е). Потомство фага может быть извлечено из клеток посредством искусственного лизиса.
7.19. Е. coli С служит нормальным хозяином фага дикого типа фХ174. У мутантного штамма Е. coli C1 поверхность клетки изменена так, что фаг фХ не может на ней адсорбироваться и соответственно не может расти и размножаться на штамме Е. coli C1. Двойной мутант фага фХ, обозначаемый символом НаНb, способен адсорбироваться на поверхности клеток штамма Е. coli C1 (равно как и на Е. coli С), инфицировать их и производить потомство. Когда клетки Е. coli С одновременно заражают фагом дикого типа и НаНb, некоторые фаговые частицы в потомстве могут инфицировать клетки Е. coli Cl однако их потомство уже лишено способности инфицировать клетки Е. coli C1. Объясните.
|
Дата добавления: 2015-12-16 | Просмотры: 841 | Нарушение авторских прав
|