|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
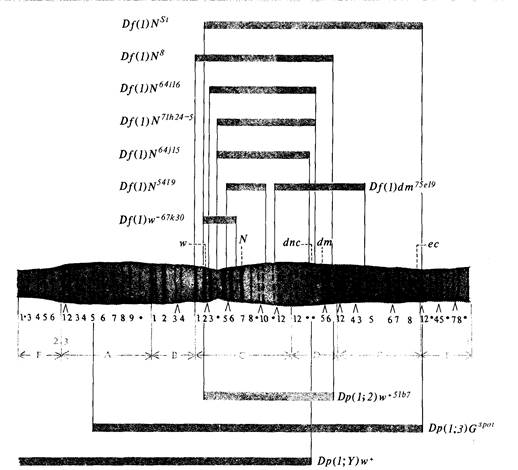
Корреляция между генетическими и цитологическими картами хромосом дрозофилыБольшая часть хромосом эукариот, в том числе и хромосомы дрозофил, характеризуется чередованием генетически активных, или эухроматиновых, участков и генетически неактивных участков, называемых гетерохроматиновыми (см. дополнение 1.1). Большинство известных генов дрозофилы расположены в эухроматиновых участках. В соматических клетках некоторых тканей дрозофилы и других видов двукрылых, в особенности в клетках слюнных желез их личинок, содержатся так назы-
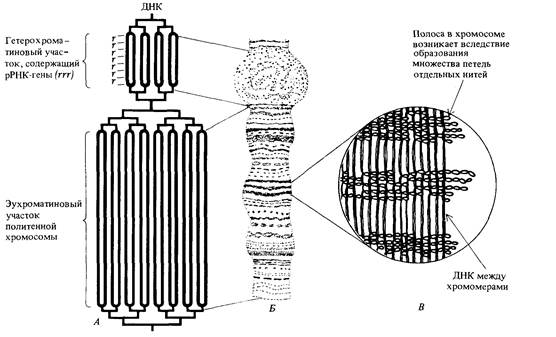
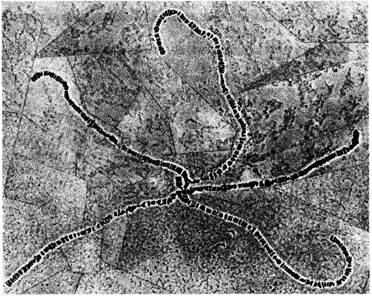
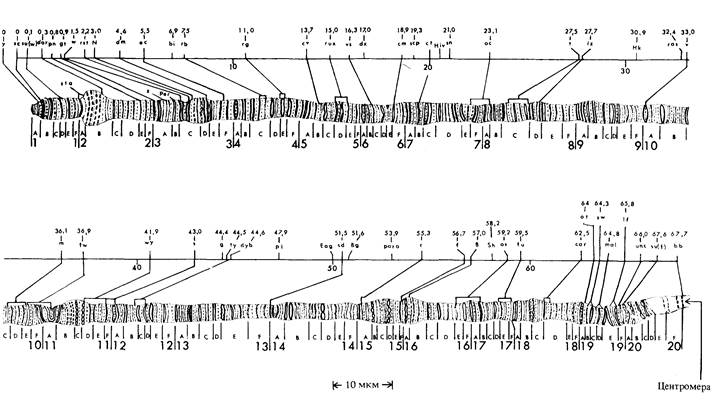
146 Организация и передача генетического материала ваемые гигантские хромосомы. Они образуются в результате множества последовательных удвоений хромосом, не сопровождающихся ядерными делениями. В зависимости от вида и стадии развития в каждом ядре каждая хромосомная нить может быть представлена более чем в 1000 копиях, и все нити обоих гомологов располагаются по всей длине в точности друг против друга, образуя политенные интерфазные хромосомы, представляющие собой подобия многожильного троса. Схематическая модель политенной Х-хромосомы изображена на рис. 5.16, а электронная микрофотография ядра клетки слюнных желез дрозофилы-на рис. 5.17. Различия в содержании белка и в степени спирализации интерфазных хромосом обусловливают их поперечную исчерчены ость, хорошо заметную на рис. 5.18. У дрозофил известны многочисленные хромосомные аберрации, при которых отдельные участки хромосом утрачиваются (делеции Df) или удваиваются (дупликации, Dp, см. гл. 21). На цитологических препаратах политенных хромосом с делециями или дупликациями концевые точки этих аберраций обычно можно определить очень точно. На рис. 5.19 изображены границы некоторых известных делеции и дупликаций в Х-хромосоме дрозофилы. Потери генетической информации, свя-
5. Геном эукариот 147
148 Организация и передача генетического материала
занные с делециями, фенотипически проявляются у гетерозигот по рецессивным мутациям. Например, у гетерозигот Df(l)N8/w, Df(l)NSt/w и Df(l)N -- 67k30/w глаза белые, что указывает на то, что в хромосомах с этими тремя нехватками ген w + отсутствует, и, следовательно, этот ген расположен в участке, общем для всех трех делеций, т. е. в полосах ЗС2-ЗС6. У гетерозиготы Df(l)N 64 · 16 /w глаза красные, таким образом, ген w + в этой хромосоме присутствует. Следовательно, ген w + должен быть расположен между левыми концами делеций Df(1)N64 i16 и Df(l)NSt или Df(l)w-- 67k30, т.е. должен находиться в полосе 3С2. Заметим, что полоса 3С2 находится внутри всех трех дупликаций, представленных на рис. 5.19. Эти участки могут быть транслоцированы во вторую аутосому, в третью аутосому и в Y-хромосому соответственно, и во всех трех случаях можно установить присутствие гена w + в геноме, что подтверждает локализацию w + в полосе 3С2. Например, самцы
150 Организация и передача генетического материала с генотипом w/Dp(1; Y)w+ обладают красными глазами, отсюда следует, что ген w+ содержится в участке Х-хромосомы, транслоцированном на Y-хромосому. Такие цитологические сопоставления были проведены для многих генов, локализованных в различных хромосомах дрозофилы, и они убедительно продемонстрировали, что генетическая карта каждой хромосомы, полученная на основе данных по рекомбинации, хорошо соответствует реальной физической структуре хромосом (рис. 5.20). Неполное количественное совпадение между расстояниями на генетической карте и реальными расстояниями в политенной хромосоме может быть следствием неравномерного растяжения различных участков хромосомы в процессе приготовления хромосомных препаратов. Возможно, однако, что некоторые участки Х-хромосомы участвуют в генетической рекомбинации чаще других и вследствие этого на генетической карте занимают больше места, чем на физической. Дата добавления: 2015-12-16 | Просмотры: 945 | Нарушение авторских прав |