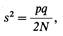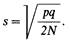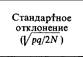|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 3. Пер. с англ.: – М.: Мир, 1988. – 336 с.126 Эволюция генетического материала нах, полученных от выросших растений, будет очень близка к 1/2, хотя может оказаться и чуть больше, и чуть меньше. Если известно число родителей в исходном поколении и частоты аллелей в нем, как это было в только что приведенном примере, то мы можем рассчитать вероятность получить в следующем поколении те или иные частоты аллелей. Для этого нам нужно знать вариансу, или дисперсию, частот аллелей в следующем поколении. Варианса служит мерой изменчивости, обнаруживаемой при сравнении различных выборок (см. приложение 3). Если имеются два аллеля с частотами p и q, причем число родителей равно N (так что число генов в исходном поколении равно 2N) то варианса (s2)частоты аллеля в следующем поколении составляет
а стандартное отклонение может быть выражено как
Эти формулы отражают обратную зависимость между величиной выборки 2N и теоретически ожидаемой изменчивостью частот аллелей. Табл. 23.6 иллюстрирует эффект дрейфа генов от одного поколения к другому в двух случаях: 1) когда p = q = 0,5 и 2) когда p = 0,3, a q = = 0,7. Для каждого случая рассматриваются три варианта эффективной численности популяции: N = 5, 50 и 500. Реально наблюдаемая частота аллеля p укладывается в интервал p ± 2 стандартных отклонений с 95% вероятностью. В малых популяциях с эффективной численностью 5 особей этот интервал ожидаемых значений p в следующем поколении лежит между 0,18 и 0,82; чем больше численность популяции, тем уже интервал ожидаемых значений частоты аллеля в следующем поколении. Заметим, что ширина этого интервала убывает с ростом эффективной численности популяции как корень квадратный из отношения эффективной численности одной популяции к эффективной численности другой.
Дата добавления: 2015-12-16 | Просмотры: 543 | Нарушение авторских прав |