|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 3. Пер. с англ.: – М.: Мир, 1988. – 336 с.23. Элементарные процессы эволюции 125
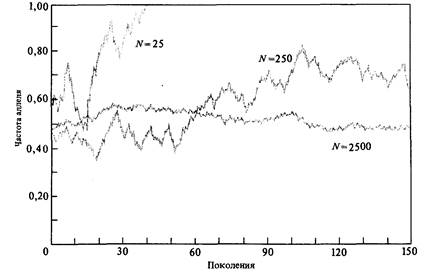
нию или снижению частоты аллеля всегда может смениться на обратную, пока частота аллеля не достигнет нуля или единицы (рис. 23.5). Если частота аллеля в одном поколении увеличилась, в следующем поколении она с равной вероятностью может либо еще больше возрасти, либо уменьшиться. Если же аллель утрачивается или «фиксируется» (т. е. если значение его частоты достигает 0 или 1), процесс прекращается. Частота аллеля уже не может более изменяться до тех пор, пока в результате мутации не появится новый аллель. Рассмотрим следующий пример. Предположим, что у нас есть множество растений гороха Pisum sativum, на котором проводил свои опыты Мендель, и что частота аллеля Y, ответственного за желтую окраску семян, равна 0,5. Такова же, естественно, и частота аллеля у, который в гомозиготном состоянии обусловливает зеленый цвет семян. Предположим также, что частоты генотипов совпадают с теоретически ожидаемыми и составляют 1/4 YY: 1/2 Yy: 1/4уу. Возьмем теперь наугад любую горошину, не обращая внимания на ее цвет, и посадим ее. Какова будет частота аллеля Y у горошин, полученных от растения, выросшего из посаженной горошины, после самоопыления? Ясно, что существует три возможности: частота аллеля Y будет равна 1, 1/2 или 0 в зависимости от генотипа посаженной горошины. С вероятностью 1/4 эта горошина обладала генотипом YY, такова же вероятность того, что ее генотип был уу; следовательно, частота аллеля Y в потомстве этой горошины с равной вероятностью принимает значение либо 0, либо 1. Предположим теперь, что мы выбрали 1000 горошин из исходной популяции и вырастили из них 1000 растений. Частота аллеля Y в гороши- Дата добавления: 2015-12-16 | Просмотры: 610 | Нарушение авторских прав |