2) Δρ = - 0,048, 3) Δρ = -0,010.
Знаменатели в выражениях для Δq и Δρ тождественны. Однако частота аллеля, против которого направлен отбор, равна q2 в случае отбора против рецессивного аллеля и р-в случае отбора против доминантного аллеля. Соответственно по мере того, как частота аллеля, против которого направлен отбор, становится все меньше и меньше, отбор против рецессивного аллеля становится все менее и менее эффективным по сравнению с отбором против доминантного аллеля. 24.5. Аллель Hbs не полностью доминантен по признаку серповидноклеточности. Что же касается его приспособленности, то он рецессивен в районах, где нет малярии, но обладает сверхдоминированием в местах, где малярия распространена. Существует множество мутаций, например мутация изогнутых крыльев Curly у дрозофилы, доминантных в отношении одного фенотипического признака (форма крыльев), но рецессивных в других отношениях (в упомянутом примере -жизнеспособность: Су -рецессивная деталь). 24.7. Аллель является рецессивной деталью; следовательно, равновесная частота
Равновесная частота рецессивной летали гораздо выше, чем доминантной, поскольку рецессивные аллели в гетерозиготном состоянии не подвержены действию отбора. 24.9. Приспособленности генотипов, приведенные в табл. 24.10, составляют: АА:( 1 — — s); Аа: 1; аа: 0. В состоянии равновесия q = = s/ (s + t) = s/(l + s) = 0,333. Следовательно, s = 0,50 и приспособленности равны: 0,50, 1 и 0. 24.11. В эксперименте 1 приспособленности равны 1, 1 и 0,50. В эксперименте 2 они равны 0,50, 1 и 1 для генотипов FF, FS, SS соответственно. Таким образом, имеет место частотно-зависимый отбор. Поскольку приспособленности гомозигот связаны с их частотой обратной зависимостью, следует ожидать возникновения устойчивого полиморфного равновесия. Приспособленности гомозигот, по-видимому, симметричны, и поэтому можно предположить, что равновесие установится при частоте аллелей, равной 0,50.

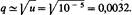 следовательно, равновесная частота
Равновесная частота рецессивной летали гораздо выше, чем доминантной, поскольку рецессивные аллели в гетерозиготном состоянии не подвержены действию отбора. 24.9. Приспособленности генотипов, приведенные в табл. 24.10, составляют: АА:( 1 — — s); Аа: 1; аа: 0. В состоянии равновесия q = = s/ (s + t) = s/(l + s) = 0,333. Следовательно, s = 0,50 и приспособленности равны: 0,50, 1 и 0. 24.11. В эксперименте 1 приспособленности равны 1, 1 и 0,50. В эксперименте 2 они равны 0,50, 1 и 1 для генотипов FF, FS, SS соответственно. Таким образом, имеет место частотно-зависимый отбор. Поскольку приспособленности гомозигот связаны с их частотой обратной зависимостью, следует ожидать возникновения устойчивого полиморфного равновесия. Приспособленности гомозигот, по-видимому, симметричны, и поэтому можно предположить, что равновесие установится при частоте аллелей, равной 0,50.
следовательно, равновесная частота
Равновесная частота рецессивной летали гораздо выше, чем доминантной, поскольку рецессивные аллели в гетерозиготном состоянии не подвержены действию отбора. 24.9. Приспособленности генотипов, приведенные в табл. 24.10, составляют: АА:( 1 — — s); Аа: 1; аа: 0. В состоянии равновесия q = = s/ (s + t) = s/(l + s) = 0,333. Следовательно, s = 0,50 и приспособленности равны: 0,50, 1 и 0. 24.11. В эксперименте 1 приспособленности равны 1, 1 и 0,50. В эксперименте 2 они равны 0,50, 1 и 1 для генотипов FF, FS, SS соответственно. Таким образом, имеет место частотно-зависимый отбор. Поскольку приспособленности гомозигот связаны с их частотой обратной зависимостью, следует ожидать возникновения устойчивого полиморфного равновесия. Приспособленности гомозигот, по-видимому, симметричны, и поэтому можно предположить, что равновесие установится при частоте аллелей, равной 0,50.