|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
С другими видами метаболизма
Общепринятым показателем, используемым в клинике в качестве индикатора состояния углеводного метаболизма, служит уровень глюкозы в крови. Её значения определяются у здорового человека сроком приёма, объёмом еды, характером питания, а также методом исследования. В среднем содержание этого моносахарида в крови в норме 3,3 - 5,5 ммоль/л. Возможно развитие двух вариантов физиологических гипергликемий: 1) алиментарная - после приёма пищи; 2) эмоциональная - следствие действия стрессоров (надпороговых раздражителей физической, химической, биологической, психогенной природы). Так как глюкоза - универсальный источний энергии, то её уровень в сосудистом русле может быстро снижаться, если его не поддерживать с помощью эндогенной формы. Основным органом, ответственным за сохранение нормогликемии, является печень, хотя определённую роль в этом играют почки, где происходит вначале фильтрация, а затем реабсорбция данного углевода в кровь. Практически все ткани способны накапливать глюкозу путем её полимеризации в гликоген, который затем при необходимости распадается (гликогенолиз). Вышеназванный процесс особенно активно протекает в гепатоцитах, так как они обладают самой высокой способностью депонировать этот гомополисахарид. А главное, в указанных клетках присутствует глюкозо-6-фосфатаза, с помощью которой цитозольный глюкозо-6-фосфат, лишаясь фосфата, приобретает способность преодолеть мембрану и выходить в сосудистое русло для поддержания гомеостаза глюкозы крови. Этот энзим регистрируется также в слизистой тонкого кишечника и, естественно, в почках (схема 2). Но запасы гликогена в организме довольно ограничены, поэтому при высокой вероятности развития гипогликемии индуцируется глюконеогенез - синтез глюкозы из продуктов неуглеводного происхождения. К ним относятся много самых различных соединений, только они должны удовлетворять следующим условиям: в их составе должно быть не менее трёх атомов углерода; если в молекуле присутствует кето-группа, необходимо, чтобы она обязательно находилась в a-положении. Следовательно, источниками глюкозы могут быть метаболиты цикла трикарбоновых кислот (ЦТК), лактат, пируват, глицерин, многие аминокислоты (гликогенные). Но высшие жирные кислоты, продукты их распада - кетоновые тела, ацетил-КоА, некоторые аминокислоты (лейцин, валин, изолейцин, фенилаланин, тирозин) (кетогенные) не способны стать участниками глюконеогенеза. Глюкоза из крови может проникнуть в нуждающиеся в ней клетки путем облегченной диффузии, в обеспечении которой большую положительную роль играет холестерин билипидного слоя плазмолемм. При гипогликемии этот способ не работает и тогда включается активный транспорт. Стимулируется локализующаяся в мембране АТФ-аза, точнее один из её вариантов - гексокиназа (глюкокиназа), с помощью её и энергии АТФ глицид уже в виде глюкозо-6-фосфата проникает в клетку. Находясь в цитоплазме, фосфопроизводное моносахарида может использоваться в следующих направлениях. Большая часть, естественно, подвергается распаду, в первую очередь, гликолизу. Этот процесс в зависимости от наличия кислорода может заканчиваться двумя продуктами: при гипоксии – лактатом, при нормоксии - пируватом, который, попадая в митохондрии, после окислительного декарбоксилирования сгорает в цитратном цикле, полностью высвобождая заложенную в глюкозе энергию. Но для клеток характерно и другое направление катаболизма данного углевода, что обычно не служит поставщиком энергии; локализуется тоже в цитоплазме и часто называется пентозофосфатным путём (ПФП). Его конечным продуктом является СО2, и кроме того окисление глюкозы сопровождается восстановлением НАДФ. Последний не идёт в митохондрии, а используется как источник водорода в различных процессах синтеза. В первую очередь, в образовании холестерина, стероидных гормонов, жёлчных кислот, а также высших жирных кислот (ВЖК) - обязательного компонента большинства липидов. Кроме того, НАДФН+Н+ - одно из важных звеньев антирадикальной защиты (АРЗ) - системы, борющейся с накоплением токсических радикалов (схема 2). Участие восстановленного НАДФ в микросомальном окислении позволяет получить тирозин, катехоламины, серотонин; этот кофермент преобразует также витамин Вс в активную форму - тетрагидрофолиевую кислоту, рибонуклеотиды в дезоксипроизводные. Метаболитом и гликолиза, и ПФП служит глицероальдегид-3-фосфат (ГА-3Ф), который после изомеризации в дигидроксиацетонфосфат с последующим восстановлением становится глицеро-1-фосфатом - субстратом в синтезе нейтральных жиров и глицерофосфатидов. Судя по названию, пентозофосфатный путь является источником различных пентоз, главная роль из которых принадлежит рибозо-5-фосфату, последний входит в состав нуклеотидов. Самые простейшие из них - мононуклеотиды - могут быть макроэргами (АТФ, ГТФ, УТФ, ЦТФ), коферментами (ФМН), внутриклеточными гормонами (циклический АМФ). Динуклеотиды обычно входят в состав ферментов в виде коэнзимов (НАД+, НАД+Ф, ФАД, КоА-SН). Нуклеиновые кислоты (РНК, ДНК) - биополимеры, химическое название которых полинуклеотиды, служат матрицами в синтезе белков, а также хранителями и передатчиками генетической информации.
Схема 2. Взаимосвязь углеводного обмена с другими видами обменов
Но не вся клеточная глюкоза обязательно подвергается распаду. Часть её активируется в УДФ-глюкозу, которая используется или в синтезе гликогена, или после преобразований в другие моносахариды и их производные - в генезе гетерополисахаридов: гиалуроновой кислоты межклеточного вещества, хондроитинсульфата хрящевой и костной тканей, гепарина - антикоагулянта, гликолипидов любых плазмолемм и специфических структур нервных клеток, гликопротеидов, служащих рецепторами в мембранах, группоспецифических веществ крови и т.д. Кроме этих, существуют и тканевые разновидности взаимоотношений обменов. В эритроцитах гликолиз протекает только в анаэробных условиях (в этих клетках полностью отсутствуют органоиды, в том числе и митохондрии - всё направлено на выполнение главной цели - как можно больше транспортировать кислорода). Конечным продуктом такого процесса (схема 3) будет лактат, он же образуется и в интенсивно работающей мышце. Механизм реакции следующий.
НОРМОКСИЯ ГИПОКСИЯ
Г-6-Ф Г-6-Ф
ГА3Ф ГА3Ф
Схема 3. Механизм образования лактата в клетке
В гликолизе происходит восстановление НАД+. Обычно высвободившийся водород в условиях нормоксии переносится в электронно-транспортную цепь (то есть вступает в процесс биологического окисления). Если же в клетке - гипоксия, это грозит накоплением протонов (НАДН+Н+), сдвигом рН (ацидозом). Чтобы предотвратить подобную опасность и осуществляется восстановление пирувата в лактат. Последнее соединение выходит из эритроцитов и мышц в кровь, доставляется в печень, где под влиянием изоэнзима ЛДГ (лактатдегидрогеназы) и окисленного НАД+ преобразуется в пируват, который, вступая в глюконеогенез, индуцирует синтез глюкозо-6-фосфата, являющегося субстратом в гликогеногенеза. При необходимости стимулируется распад гомополисахарида (гликогена) и глюкоза попадает в сосудистое русло, откуда в мышцы, где вновь может подвергнуться гликолизу (эта циркуляция называется циклом Кори). Учитывая, что лактатдегидрогеназа обладает тканевой специфичностью (в миоцитах работает на образование молочной кислоты, а в гепатоцитах - наоборот, на окисление её до ПВК), можно предположить определённые специфические особенности в их строении, и отсюда в их физических свойствах. А это, как известно, используется в дифференциальной диагностике болезней миокарда и печени.
Дата добавления: 2015-12-16 | Просмотры: 674 | Нарушение авторских прав |





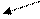
 НАД+ НАД+
НАД+ НАД+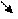
 ЭТЦ
ЭТЦ

 НАДН+Н+ Н2О НАДН+Н+ ЭТЦ
НАДН+Н+ Н2О НАДН+Н+ ЭТЦ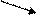 О2
О2 1,3 ДФГ 1,3 ДФГ
1,3 ДФГ 1,3 ДФГ


 ПВК ПВК
ПВК ПВК
 ЛДГ
ЛДГ НАД+
НАД+ лактат кровь
лактат кровь