|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Клеточная мембрана. Мембрана клетки представляет собой мозаику из липидов и белков, её толщина около 6-8 нанонеметров (нм)Мембрана клетки представляет собой мозаику из липидов и белков, её толщина около 6-8 нанонеметров (нм). Липиды клеточной мембраны по большей части относятся к фосфолипидам, в молекулах которых есть полярная, т.е. несущая электрический заряд, головка и два неполярных хвоста, построенных из атомов углерода и водорода (Рис. 1.3). Такие молекулы плохо растворяются в воде – полярном растворителе: растворению мешают их неполярные хвосты. Поэтому в воде фосфолипиды образуют мицеллы – микроскопические капельки, внутри которых прячутся неполярные хвосты молекул, а полярные головки обращены наружу – к воде. Когда концентрация фосфолипидов высока, мицеллы соединяются друг с другом так, что образуется двойной или бимолекулярный липидный слой, внутрь которого обращены гидрофобные хвосты, а наружную поверхность представляют гидрофильные полярные головки.
В бимолекулярном липидном слое находятся мембранные белки, молекулы которых значительно крупнее, чем у фосфолипидов, и зачастую свёрнуты наподобие клубка, образуя довольно объёмную структуру: она называется глобулой. Неполярные части белковых молекул обычно погружены вовнутрь, а полярные выступают над мембранной поверхностью, как с наружной, так и с внутренней стороны. Есть и такие крупные белковые молекулы, которые пронизывают мембрану насквозь. Их принято называть интегральными белками в отличие от остальных, названных периферическими. Погружённые в жидкую плёнку бимолекулярного слоя липидов белки способны медленно перемещаться из одного участка в другой; используя метафору, о мембране можно сказать так: это липидное море, в котором, как айсберги, плавают белки. Каким путём могут пройти через мембрану необходимые клетке вещества, как удаляются продукты её жизнедеятельности? Жирорастворимые вещества, естественно, растворяются и в липидах мембраны и поэтому могут довольно легко пройти через неё путём обыкновенной диффузии. Так же легко диффундируют через липидную часть мембраны растворимые в жидкостях газы, например кислород и углекислый газ. Но растворённые в воде молекулы (обычно несущие электрический заряд), ионы и крупномолекулярные соединения способны пройти через мембрану только с помощью специальных транспортных белков, среди которых различают каналы и насосы. Каналы – это трубчатые белки, они имеют заполненную водой пору, через которую по концентрационному или электрическому градиенту проходит тот или иной ион либо молекула. Такой транспорт называют пассивным, поскольку он не требует расхода энергии специально для переноса. Иное дело, если ионы или молекулы понадобится перенести против концентрационного или электрического градиента: в этом случае понадобится энергия. Такой транспорт назван активным и его осуществляют белки-насосы, которые используют энергию аденозинтрифосфорной кислоты (АТФ). Многие мембранные белки действуют в качестве ферментов: они ускоряют биохимические реакции в самой мембране и возле её поверхностей. Ферменты высоко специфичны, т.е. каждый из них контролирует только одну биохимическую операцию. В связи с этим каждой клетке приходится иметь не одну сотню различных ферментов, как механику, вынужденному носить с собой набор различных гаечных ключей. Клеточные рецепторы – ещё одна разновидность мембранных белков, Они выступают над наружной поверхностью мембраны и в этой части своей молекулы имеют участки, специфически связывающие строго определённые вещества: нейромедиаторы, гормоны или иные биологически активные соединения. Прикрепление такого вещества к рецептору влияет на деятельность клетки, например, изменяет проницаемость её мембраны или скорость обменных реакций внутри клетки. Некоторые белки нужны для сохранения формы клетки и субклеточных структур, для объединения клеток друг с другом – такие белки называют структурными. Все белки, независимо от выполняемой функции, со временем разрушаются, а на смену им синтезируются новые белковые молекулы. Мембранные белки определяют специфическое поведение той или иной клетки, именно от них зависит: какие именно вещества, и в каком количестве смогут войти в клетку или покинуть её. Это обстоятельство важно не только для отдельно взятой клетки, но и для межклеточных отношений, т.е. для межклеточной сигнализации. Передача сигнала от клетки к клетке возможна только двумя способами: проведением электрического тока или использованием специальных химических веществ в качестве курьеров для передачи информации. И в том, и в другом случае клеточные мембраны должны избирательно изменять свою проницаемость, избирательно регулировать характер биохимических реакций, избирательно связываться с определёнными веществами и т.д. По-разному решая все эти проблемы выбора, клетки демонстрируют свою индивидуальность только благодаря индивидуальному подбору белков. Как уже было сказано, органеллы клетки имеют собственные мембраны. К этому следует добавить, что они во многом напоминают плазматическую мембрану клетки по своему строению и функциональной организации. Ядро клетки Ещё в XIX веке в ядре были обнаружены хромосомы – интенсивно окрашивающиеся тельца, которые регулярно появляются во время клеточного деления или митоза. Позже стало известно, что хромосомы – это плотно упакованные в ядерные белки двойные спирали дезоксирибонуклеиновой кислоты (ДНК). У каждого вида животных обнаруживается постоянное число хромосом определённой формы и величины. У человека есть 23 пары гомологичных, т.е. соответствующих друг другу по определяющим признакам, хромосом. Одна из хромосом каждой пары унаследована от отца, другая – от матери. Поскольку все клетки организма происходят от одной оплодотворённой яйцеклетки, они имеют совершенно одинаковый набор хромосом вне зависимости от принадлежности клетки к той или другой ткани.
В определённой последовательности этих нуклеотидов заключена вся генетическая информация ДНК: комбинация трёх следующих друг за другом нуклеотидов (она называется триплет или кодон) означает выбор одной из 20 существующих аминокислот для включения её в молекулу вновь синтезируемого белка. Каждый из четырёх нуклеотидов можно представить, как "букву генетического языка", а триплет – как кодовое слово, из которых составляется более или менее длинная фраза. Она содержит полную инструкцию для синтеза какого-либо конкретного белка, а участок хромосомы, на котором записана такая информация, получил название – ген. Каждый ген содержит также знаки препинания в виде т.н. инициирующих и терминирующих кодонов, которые определяют начало и конец считывания генетической информации. Существуют специальные механизмы экспрессии генов, т.е. востребования генетической информации. При возникновении потребности в каком-либо белке происходит деконденсация хромосомы, т.е. разуплотнение в том локусе (участке), где находится нужный ген. Двойная спираль ДНК в этом месте расходится, чтобы предоставить возможность для синтеза молекулы информационной рибонуклеиновой кислоты (иРНК). Этот синтез – не что иное, как переписывание генетической информации (транскрипция), причём транскрипция контролируется специальным ферментом. Когда же транскрипция завершится, разошедшиеся для неё нити ДНК вновь соединятся, как застёжка на молнии. Образующаяся иРНК отличается от ДНК тем, что имеет лишь одну цепь, синтезируемую на нити ДНК, как на матрице. Кроме того, вместо D-2-дезоксирибозы она содержит другой углевод – рибозу, а вместо тимина другой нуклеотид – урацил. При синтезе иРНК аденин подстраивается напротив тимина ДНК, а заменяющий тимин урацил – напротив аденина ДНК. Цитозин располагается напротив гуанина и наоборот, а в результате в точности воспроизводится генетический код ДНК. Затем происходит редакция считанного материала, которая заключается в вырезании не кодирующих последовательность аминокислот участков генома (они были нужны лишь для транскрипции). Полученная в итоге молекула иРНК содержит всю информацию о синтезе нужного белка, сам же синтез произойдёт в другом месте, после того, как молекула иРНК уйдёт из ядра сквозь пору ядерной оболочки. Таким образом, функция клеточного ядра связана с хранением генетической информации. При появлении запроса на какую-либо её часть информация копируется с ДНК на иРНК, которая уносит полученную инструкцию для синтеза белка к месту такого синтеза. Другая сторона деятельности клеточного ядра связана с его делением путём митоза. Этот вопрос здесь не рассматривается, поскольку сформированные нервные клетки не делятся. Описанный выше механизм извлечения генетической информации используется всегда, когда требуется заменить изношенные белковые молекулы, синтезировать вновь понадобившиеся ферменты, увеличить количество каких-либо рецепторов или структурных белков. Отдельные виды генов кодируют, например, образование больших молекул-предшественниц, из которых затем формируются т.н. нейропептиды, используемые нервными клетками для передачи информации другим клеткам. Все эти процессы усиливаются при формировании новых образцов поведения, т.е. при научении, когда образуются энграммы памяти (следы). Не удивительно, что в нервной ткани обнаружено около 200 000 разнообразных молекул иРНК, что в 10-20 раз больше, чем, например в печени или почках. С обилием используемой в нервной ткани генетической информации отчасти связано и многообразие клеточных типов нейронов. Рибосомы Маленькие тельца диаметром около 20-22 нм, не имеют собственной мембраны и содержат особую разновидность РНК, которая называется рибосомальной. После выхода из ядра молекулы иРНК к ней присоединяется несколько рибосом, в результате чего образуется полисома, на которой происходит синтез белка (рис.1.5). Синтез заключается в том, чтобы собрать аминокислоты в определённой последовательности, закодированной триплетами иРНК. Каждую аминокислоту представляет для синтеза специфическая транспортная РНК. На одном конце её молекулы есть участок для соединения с определённой аминокислотой, а на другом – антикодон, т.е. три нуклеотида, комплементарных (взаимно дополняющих) соответствующему кодону иРНК.
Некоторая часть рибосом и полисом находится поблизости от ядра в цитозоле – их называют свободными. Другая часть рибосом прикреплена к мембране эндоплазматического ретикулума. На свободных рибосомах обычно синтезируются белки для цитозоля, микротрубочек, микрофиламентов, т.е. для внутреннего использования, тогда как на связанных с эндоплазматическим ретикулумом рибосомах образуются белки, предназначенные для выделения из клетки, например, нейропептидные медиаторы. Дата добавления: 2015-12-15 | Просмотры: 1132 | Нарушение авторских прав |
 Именно так и выглядит липидная основа клеточной мембраны. Кроме представляющего фосфолипиды фосфатидилхолина в ней есть гликолипиды. Они обычно расположены на наружной поверхности мембраны так, что выступающие углеводные части молекул образуют надмембранный слой. Ещё один компонент мембраны – холестерин, напротив, находится во внутреннем слое мембраны и играет роль регулятора агрегатного состояния липидной части мембраны: плотную мембрану он разжижает, а жидкую – уплотняет. Нормальное состояние мембраны – это жидкая плёнка определённой вязкости, примерно соответствующей вязкости оливкового масла.
Именно так и выглядит липидная основа клеточной мембраны. Кроме представляющего фосфолипиды фосфатидилхолина в ней есть гликолипиды. Они обычно расположены на наружной поверхности мембраны так, что выступающие углеводные части молекул образуют надмембранный слой. Ещё один компонент мембраны – холестерин, напротив, находится во внутреннем слое мембраны и играет роль регулятора агрегатного состояния липидной части мембраны: плотную мембрану он разжижает, а жидкую – уплотняет. Нормальное состояние мембраны – это жидкая плёнка определённой вязкости, примерно соответствующей вязкости оливкового масла. ДНК – это высокомолекулярный полимер, образованный соединёнными друг с другом нуклеотидами, каждый из которых состоит из молекулы углевода D-2-дезоксирибозы, остатка фосфорной кислоты и одного из четырёх азотистых оснований: аденина, гуанина, цитозина и тимина. В 1953 году Джеймс Уотсон и Фрэнсис Крик (Watson J. Crick F.) построили модель ДНК, которая состоит из двух длинных цепей, скрученных в виде спирали; при этом обращённые внутрь азотистые основания попарно соединены друг с другом водородной связью, причём аденин всегда соединён с тимином, а гуанин – с цитозином (рис. 1.4).
ДНК – это высокомолекулярный полимер, образованный соединёнными друг с другом нуклеотидами, каждый из которых состоит из молекулы углевода D-2-дезоксирибозы, остатка фосфорной кислоты и одного из четырёх азотистых оснований: аденина, гуанина, цитозина и тимина. В 1953 году Джеймс Уотсон и Фрэнсис Крик (Watson J. Crick F.) построили модель ДНК, которая состоит из двух длинных цепей, скрученных в виде спирали; при этом обращённые внутрь азотистые основания попарно соединены друг с другом водородной связью, причём аденин всегда соединён с тимином, а гуанин – с цитозином (рис. 1.4).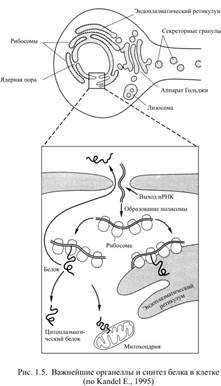 Перенос информации из генетического кода иРНК в аминокислотную последовательность белковой молекулы называется трансляцией. При этом рибосома перемещается по нити иРНК и одновременно с этим одна за другой, в нужной последовательности соединяются аминокислоты. Представив требующуюся аминокислоту, транспортная РНК отделяется от неё и снова соединяется с такой же аминокислотой, найдя её в цитоплазме.
Перенос информации из генетического кода иРНК в аминокислотную последовательность белковой молекулы называется трансляцией. При этом рибосома перемещается по нити иРНК и одновременно с этим одна за другой, в нужной последовательности соединяются аминокислоты. Представив требующуюся аминокислоту, транспортная РНК отделяется от неё и снова соединяется с такой же аминокислотой, найдя её в цитоплазме.